

{kind=link}
{kind=link}
野生地果的光合日进程及其影响因子
[王鑫a  , 笪文怡
, 笪文怡b , 唐娅b, * ]
, 笪文怡]
|
|


作者简介:王 鑫(1994—),男,贵州安顺人,学士,研究方向为植物生理学,E-mail:794302478@qq.com。
以地果为实验材料,利用LI-6400XT光合测定系统对自然条件下地果的生理指标进行了测定,并用SPSS19.0软件分析了地果的光合生理特征参数和环境因子日变化的相关性,为实现地果的人工种植提供理论依据。结果表明:地果叶片的最大净光合速率(CO2)约为15.5 μmol·m-2·s-1;净光合速率和蒸腾速率日变化都呈双峰型曲线,影响净光合速率的主要环境因素有光合有效辐射和大气相对湿度。净光合速率和大气相对湿度呈正相关与胞间CO2浓度、气孔导度呈负相关。
地果(Ficus tikoua Bur.)为桑科菠萝蜜亚科、榕属无花果亚属的落叶匍匐木质藤本植物, 俗称地石榴, 常生长在草坡、荒地或岩石缝中, 主要分布在广西、云南、四川等地[1], 是我国少数民族常用的中药材[2]。其根的药用价值有泄泻、黄疸、瘰疬、痔疮、遗精、清热利湿[3]; 茎叶用于风热咳嗽、水肿、经闭、痢疾、带下病; 花用于遗精、滑精; 果实可以祛风除湿, 治疗咽喉痛、清热散寒等[4]。除此之外, 地果还是很好的园林观赏植物, 可以防沙固土, 同时适合套栽在果园里, 使传统的单季果园成为双季果园[5]。综上所述, 地果是一种集食用、药用、绿化、观赏于一体的植物, 具有较好的经济效益。本文以野生地果为研究材料, 研究了野生地果光合日变化及其环境因子日变化的相关性, 为人工种植提供理论指导, 同时光合作用也是评价植物生产能力的重要方法[6]。
野生地果主要分布在广西、四川、贵州、云南等地, 为匍匐木质藤本, 茎上生长细长的不定根, 节膨大; 幼枝表面被有短刺毛, 背面沿脉有细毛[7]; 榕果成对或簇生于匍匐茎上, 多数埋在土中, 球形至卵球形, 直径1~2 cm, 基部收缩成狭柄, 成熟时深红色, 表面有圆形瘤点, 基生苞片3, 细小; 花期5— 6月, 果期7月[8]。
试验地位于四川省南充市西华师范大学内废弃的铁路边, 地理坐标30° 49'N, 106° 04'E, 海拔300 m, 土壤以紫色土为主[9]。试验地土壤为砂土, 年日照时数约1 538.8 h, 最热为8月, 月日照时数为218.1 h[10]。
选择晴天, 采用L1-6400XT便携式光合测量系统, 观察自然条件下地果的净光合速率日变化及环境因子日变化的关系。随机选长势相同的3株地果, 8:00— 18:00, 每隔1 h对选取的3株地果各测量1次, 每次测量取同一株的3片叶子(由顶端往下数的第3、4、5片叶子), 3株测一个轮回。测定指标包括叶片净光合速率(Photosynthetic rate, Pn)、气孔导度(Conductance to H2O, Gs)、蒸腾速率(Transpiration rate, Tr)、胞间CO2浓度(Intercellular CO2 concentration, Ci)、光合有效辐射(External PAR, PAR)、大气温度(Temperature in sample cell, Ta)、空气相对湿度(Relative humidity in the sample cell, RH)和大气CO2浓度(Sample cell CO2, Ca)等指标。
利用Excel对试验所得数据进行整理, 光合特征参数相关性分析以及其他统计分析均采用SPSS 19.0软件, 图表处理采用Excel。
从图1中a可知, 光合有效辐射日变化呈现“ 单峰” 型曲线, 在中午13:00达到最大值, 约1 822.2 μ mol· m-2· s-1, 然后开始下降, 到下午18:00时光合有效辐射大约为411 μ mol· m-2· s-1。大气CO2浓度从早上8:00到中午12:00急剧下降, 之后趋于稳定。光合有效辐射和大气CO2浓度没有相关性。
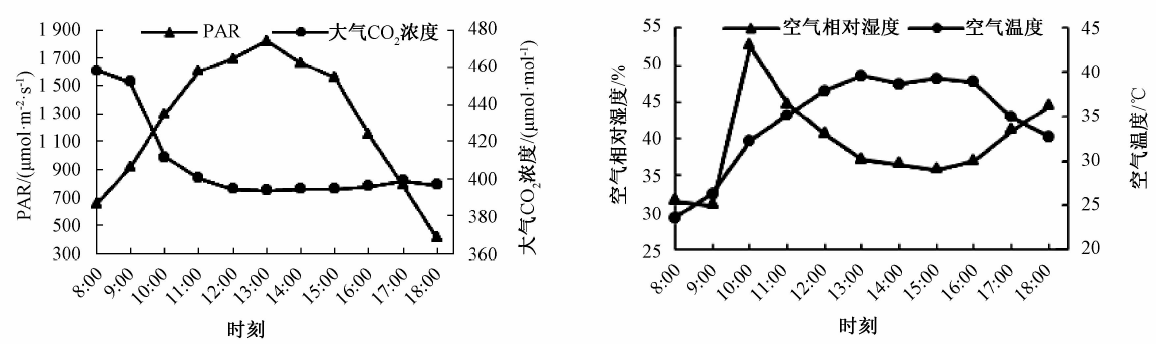 | 图1 环境因子的日变化 |
从图1中b可知, 大气温度日变化呈“ 单峰” 型曲线, 于中午13:00时达到全天的最高温度, 约40 ℃。空气相对湿度早晚较高, 由于早上露水较重, 所以随温度的上升, 8:00— 10:00空气湿度急剧上升, 在10:00左右达到最高; 然后开始下降, 下午15:00出现最低点, 约36%, 之后又大幅回升。空气相对湿度和空气温度呈负相关。
由图2中a可知, 野生地果在晴天日照条件下, 净光合速率日变化呈“ 双峰” 型曲线, 出现“ 午休” 的现象, 第1个峰值出现在上午10:00左右, 且峰值较高, 约为15.51 μ mol· m-2· s-1, 第2个峰值出现在下午15:00左右, 约为13.14 μ mol· m-2· s-1。光合有效辐射日变化呈“ 单峰” 型曲线, 达到全天最高时净光合速率处在“ 峰谷” 。净光合速率和光合有效辐射日变化呈正相关(r=0.613* )。
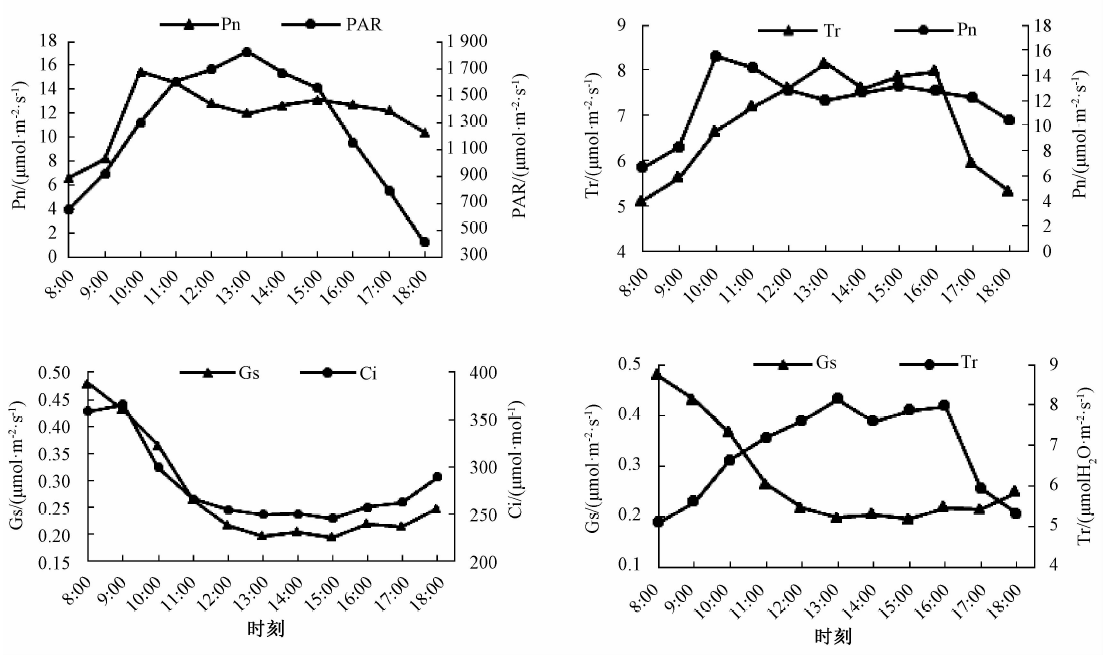 | 图2 地果叶片的光合参数日变化 |
由图2中b可知, 蒸腾速率日变化呈双峰型曲线, 第1个峰值大约出现在中午13:00, 约为8.15 mmol· m-2· s-1, 同时也是净光合速率日变化出现峰谷的时间; 第2个峰值出现在下午16:00左右, 大约为7.97 mmol· m-2· s-1。植物通过蒸腾作用为光合作用提供所需的水分、营养物质, 并调节叶面温度等, 与植物的净光合速率密切相关。
由图2中c可知, 野生地果的胞间CO2浓度日变化从早上开始呈现下降趋势, 在早上最高, 约为366.5 μ mol· mol-1, 到下午15:00降到最低, 约为245.5 μ mol· mol-1, 随后出现小幅的回升。气孔导度日变化同样在早上最高, 日变化也呈下降趋势, 下午15:00降到最低, 约0.193 mol· m-2· s-1, 胞间CO2浓度与气孔导度呈明显的正相关(r=0.962* * )。
由图2中d可知, 野生地果的气孔导度在早上最高, 约为0.479 mol· m-2· s-1, 随日进程呈现下降趋势, 到下午15:00达到最低, 约为0.193 mol· m-2· s-1, 之后略微回升。蒸腾速率日变化呈现双峰型曲线, 气孔导度和蒸腾速率日变化关系呈负相关(r=-0.707* )。气孔导度受环境因子的影响较大, 与空气温度呈负相关(r=-0.951* * ), 与大气CO2浓度呈正相关(r=0.956* * ), 适宜的大气CO2浓度和温度有利于气孔的打开, 气孔阻力降低, 气孔导度增大。
光合作用是植物十分复杂的生理过程, 研究结果(表1)表明, 一天中野生地果成熟叶片Pn与Tr呈正相关, 与Ca、Ci、Gs呈负相关。
| 表1 光合特征参数和环境因子的相关性 |
在植物生长发育时期, 净光合速率与环境因子的关联较紧密。通过对野生地果净光合速率日变化与环境因子日变化的相关性分析发现, 野生地果成熟叶片的净光合速率日变化呈双峰型曲线, 第1峰值出现在上午10:00, 第2峰值则出现在下午15:00, 此时地果的光合能力与上午10:00时相比有小幅下降, 即地果出现光合速率的午间降低现象(光合午休), 这是C3植物在高温天气时常有的普遍现象[11]。不同的植物导致光合午休的原因很可能是不相同的, 主要有气孔限制因素和非气孔限制因素[12]。舒英杰等[13]对茄子光合“ 午休” 现象的研究表明, 光合“ 午休” 时气孔导度、胞间CO2浓度、大气CO2浓度均下降, 而光合有效辐射此时几乎处于最大值。茄子光合午休是由气孔因素引起的。柯世省等[14]对浙江天台山茶树的光合日变化及光响应的研究发现, 在初夏晴天, 上年越冬叶片的光合速率、表观量子效率、羧化效率和饱和光强比展叶约16 d的一年生叶高, CO2补偿点和光补偿点比一年生叶低, 两者的净光合速率日进程曲线均为双峰型, 午间胞间CO2浓度上升表明, 此时净光合速率下降主要受非气孔限制因素的影响; 一年生叶蒸腾速率高于二年生叶, 它们的日进程曲线为单峰型, 午间最高, 而气孔阻力午间最低。光合作用是植物十分复杂的生理过程, 叶片光合速率与自身因素和环境因素密切相关[15]。该研究表明, 自然条件下生长的地果成熟叶片的净光合速率和蒸腾速率呈正相关, 净光合速率日变化和胞间CO2浓度、气孔导度呈负相关。当地果出现光合午休时, 胞间CO2浓度、气孔导度和大气CO2浓度也下降, 光合有效辐射最高。随后, 胞间CO2浓度和气孔导度小幅回升, 净光合速率也回升, 这表明野生地果叶片净光合速率午间降低主要受气孔导度等因素的影响。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|