{kind=link}
{kind=link}
{kind=link}
珍稀濒危植物天目铁木群落结构及物种多样性
[罗远1  , 吴世斌
, 吴世斌2 , 库伟鹏3 , 杨淑贞2 , 吴家森3, * ]
, 吴世斌]
|
|


作者简介:罗远(1975—),浙江杭州人,助理工程师,本科,从事生物多样性保护工作,E-mail:2630456134@qq.com
以浙江天目山国家级自然保护区的天目铁木 Ostrya rehderiana群落为对象,采用标准样地法研究了该群落植物种类、数量特征、物种多样性和群落的垂直和径阶结构。结果表明:该群落资源丰富,共有维管束植物43科67属73种。天目铁木群落垂直结构明显,可分为乔木层、灌木层和草本层,其中乔木层优势树种为天目铁木和马尾松 Pinus massoniana,灌木层优势种为隔药柃 Eurya muricata、苎麻 Boehmeria nivea,草本层优势种为水竹 Phyllostachys heteroclada、水蓼 Polygonum hydropiper。群落中的生活型以高位芽植物数量最多,占32.9%。群落中灌木层植物最为丰富,多样性指数Shannon-Weiner、优势度指数Simpson和均匀度指数Pielou均大于草本层和乔木层。天目铁木单一树种的垂直和径阶结构均为偏峰型,表现为衰退型群落特征。幼苗个体缺乏和人为干扰是导致其濒危的重要原因,因此,建议在目前封山育林保护基础上,加强天目铁木高效繁育技术研究,并适当进行人工抚育,以保证其种群的正常更新。
森林群落的物种多样性和垂直、径阶结构反映了森林群落的形成、稳定性及演替规律[1, 2]。开展濒危植物群落结构及物种多样性的研究, 可以了解种群的数量动态及演变趋势, 揭示珍稀植物的濒危过程及濒危机理[3], 为濒危植物的就地和迁地保护提供理论基础。天目铁木Ostrya rehderiana, 桦木科Betulaceae铁木属Ostrya植物, 为国家Ⅰ 级重点保护野生植物, 仅分布于浙江省杭州市临安区天目山镇大有村。前人已对天目铁木的花及花粉形态[4, 5, 6]、种子特性及繁殖[7, 8]、遗传多样性及芽苗诱导[9, 10, 11]、光合及蒸腾特性[12]、木材力学性质[13]等进行了研究, 但研究对象仅为个体或单株材料。珍稀植物的生长与周围的乔木、灌木、草本等森林植物和环境发生着各种协调、竞争的相互关系, 因此, 开展天目铁木乔木层树种的树高、径阶结构和乔、灌、草植物的多样性等群体特征的研究显得尤为重要。本文通过对天目铁木群落的区系组成、结构和演替规律等进行研究, 可为就地和迁地保护提供理论基础。
研究区位于浙江省杭州市临安区天目山镇, 天目铁木群落的中心地理位置为30° 17'N, 119° 27'E。属北亚热带季风性气候, 年均气温为14.8 ℃, 最冷月(1月)平均气温3.4 ℃, 最热月(7月)气温28.1 ℃。年降水量1 390 mm, 年日照时数1 550 h, 无霜期235 d。天目铁木群落位于海拔250 m的山麓林缘处, 南坡, 坡度15° , 土壤为发育于凝灰岩的红壤土类。
1.2.1 调查方法
2017年8月上旬对天目铁木群落进行全面调查。采用典型抽样法, 以天目铁木分布中心为起点, 沿坡向在山体上坡至下坡地段, 布设20 m× 20 m的样地3个, 再将样地划分成16个5 m× 5 m小样方。利用胸径尺调查胸径≥ 5.0 cm乔木层的不同树种, 同时采用测高仪法调查株高; 全面调查灌木层、草本层的植物种类、株数和盖度[2]。
1.2.2 垂直结构和径阶结构划分
根据天目铁木群落的株高特点, 将乔木层的垂直结构划分为3个等级, 每个等级间隔5 m, 株高分别为3~< 8 m, 8~< 13 m, 13~< 18 m。乔木层径阶结构划分为7个等级, 等级间相隔5 cm, 分别为5~< 10 cm, 10~< 15 cm, 15~< 20 cm, 20~< 25 cm, 25~< 30 cm, 30~< 35 cm, ≥ 35 cm。
1.2.3 数据分析
不同植物的重要值、多样性指数Shannon-Weiner、优势度指数Simpson和均匀度指数Pielou计算按郑昌荣等[14]的方法进行; 植物生活型谱统计参考高贤明[15]的方法进行; 种子植物地理成分根据吴征镒[16]的方法进行统计分析。
天目铁木群落中共有维管植物43科67属73种。其中蕨类植物2科2属2种, 裸子植物2科2属2种, 被子植物39科61属69种。种类数量占优势的科由大到小依次为禾本科(7属7种), 菊科(7属7种), 豆科(5属5种), 百合科Liliaceae(3属3种), 樟科Lauraceae(2属3种), 蔷薇科(1属3种)。含有单种的属占绝对优势, 有61个, 占总属数的91.0%。从以上可知, 天目铁木群落科属组成复杂, 植物种类丰富。
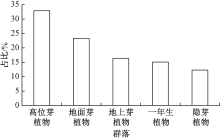
如图1所示:天目铁木群落中, 不同生活型植物种类所占比例依次为高位芽植物(32.9%)> 地面芽植物(23.3%)> 地上芽植物(16.4%)> 一年生植物(15.1%)> 隐芽植物(12.3%)。
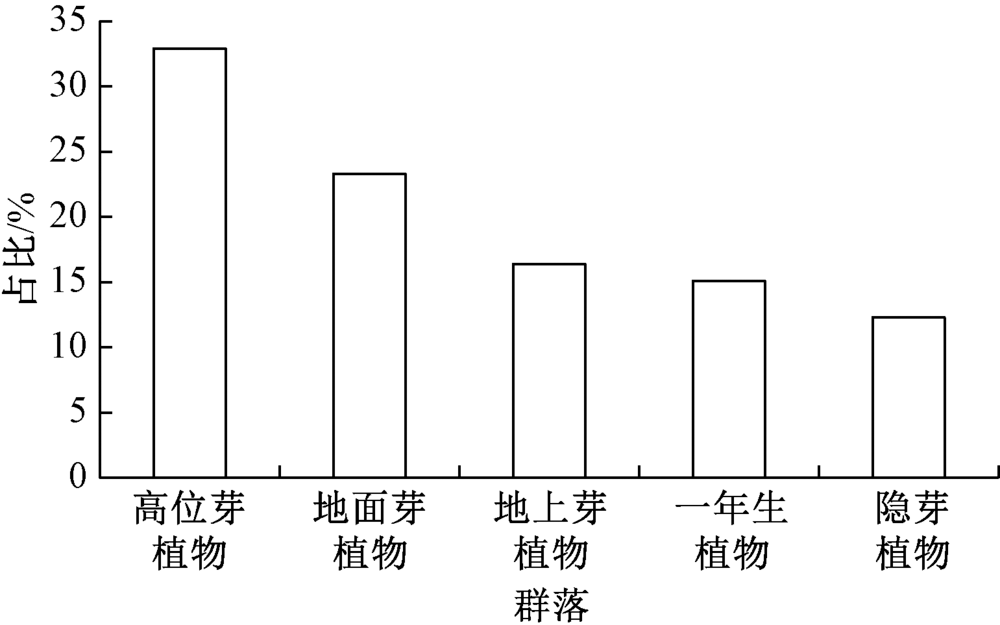 | 图1 天目铁木群落不同生活型谱 |
从表1可知:天目铁木群落中种子植物属以泛热带分布的类型有15属, 占总属数的22.4%, 如菝葜属Smilax, 紫金牛属Ardisia和冬青属Ilex等。其他依次为广布类型有11属, 占总属数的16.4%, 如悬钩子属Rubus, 飞蓬属Erigeron和商陆属Phytolacca等。东亚分布类型有9属, 占总属数的13.4%, 如山麦冬属Liriope, 木通属Akebia和沿阶草属Ophiopogon等。北温带, 东亚和北美洲间断2个分布类型均为7属, 分别占总属数的10.4%。该群落中没有植物分布的类型区有旧世界温带, 温带亚洲, 地中海地区、西亚至中亚和中亚4个。
| 表1 天目铁木群落种子植物属的分布区 |
天目铁木群落具有完整的垂直结构, 具有乔木层、灌木层、草本层和层间植物。乔木层共有7种植物(表2), 重要值最大的2种植物分别为马尾松Pinus massoniana(38.834 7)和天目铁木(37.996 6), 是该群落的主要建群树种, 控制着群落的组成、结构和生境, 其余5种乔木依次为化香Platycarya strobilacea、杉木Cunninghamia lanceolata、青冈Cyclobalanopsis glauca、枫香Liquidambar formosana、浙江樟Cinnamomum chekiangense。从树种组成可知, 该群落为马尾松-天目铁木常绿、落叶针阔混交林。
| 表2 天目铁木群落主要植物的重要值 |
灌木层发达, 生长良好, 平均高2.2 m, 盖度45%。灌木中排名前10种优势植物的重要值如表2所示。重要值大小依次分别为:隔药柃Eurya muricata、苎麻Boehmeria nivea、山莓Rubus corchorifolius、茶Camellia sinensis、浙江樟Cinnamomum chekiangense、蓬蘽Rubus hirsutus、楤木Aralia chinensis、山胡椒Lindera glauca、大青Clerodendrum cyrtophyllum、雀梅Sageretia thea。
草本层平均高0.4 m, 盖度20%。排名前10位的优势草本植物排序分别为水竹Phyllostachys heteroclada、水蓼Polygonum hydropiper、微糙三脉紫菀Aster ageratoides var.scaberulus、栗褐薹草Carex brunnea、短穗竹Semiarundinaria densiflora、圆果雀稗Paspalum orbiculare、山麦冬Liriope spicata、虎杖Polygonum cuspidata、黑足鳞毛蕨Dryopteris fuscipes、美洲商陆Phytolacca americana。
天目铁木群落植物物种多样性指数如表3所示, 从表中可知Shannon-Wiener多样性指数、Simpson优势度指数和Pielou均匀度指数在各层的排列顺序均表现为灌木层> 草本层> 乔木层。灌木层物种数量相对乔木层更为丰富, 主要原因是该层树种除了灌木外, 还包括了乔木层树种中的幼苗、幼树等, 如褐叶青冈、尾叶冬青。
| 表3 群落乔木、灌木和草本层物种多样性 |
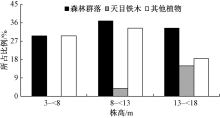
天目铁木群落的垂直结构如图2所示。整个森林群落和其他树种(除天目铁木外)的乔木层树高在3~18 m, 随着树高的增长, 植物个体所占比例先增加而后下降, 呈现近正态分布的特点; 而天目铁木树高分布呈现明显的偏峰型结构, 树高大于13 m的天目铁木占整个群落的14.8%, 而小于8 m的个体则为0。
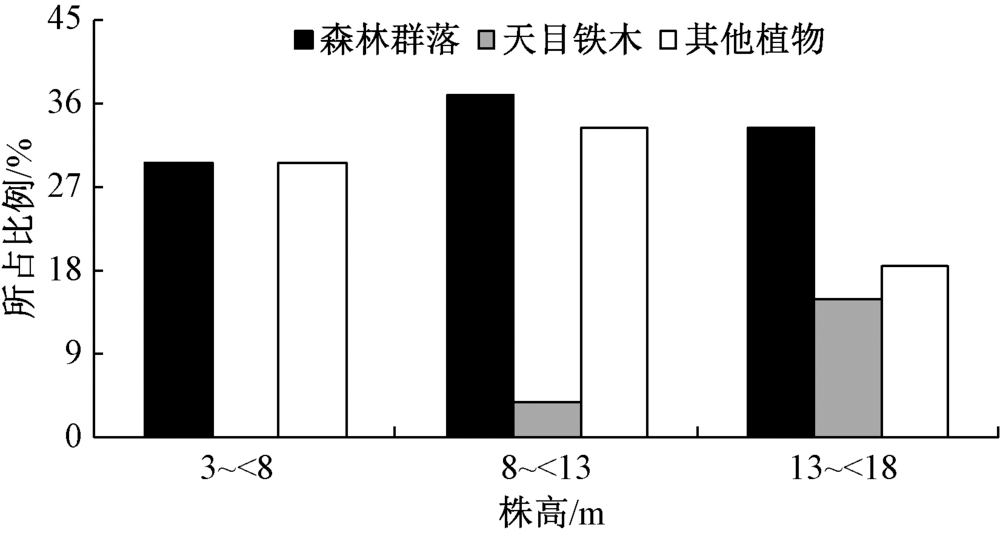 | 图2 乔木层树种的树高分布 |
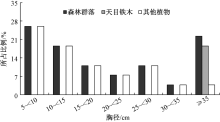
与群落的垂直结构相似, 天目铁木的径阶结构呈现出明显的偏峰型(图3), 所有个体的胸径均大于35.0 cm, 占整个乔木层的18.5%; 而其他植物的径阶分布为“ 逆J-字型” , 其形状与倒逆放置的“ J” 字型相似, 随着径阶的增大, 其个体数量相对减少, 即表现为小径阶的大峰和大径阶的小峰; 整个植物群落的径阶结构则呈现为“ V” 字型, 即随着径阶的增大, 其个体数量先减少, 当径阶大于35.0 cm时, 其个体数量又显著增加, 5 cm~10 cm和≥ 35 cm的个体数分别占整个乔木层的25.9%和22.2%。
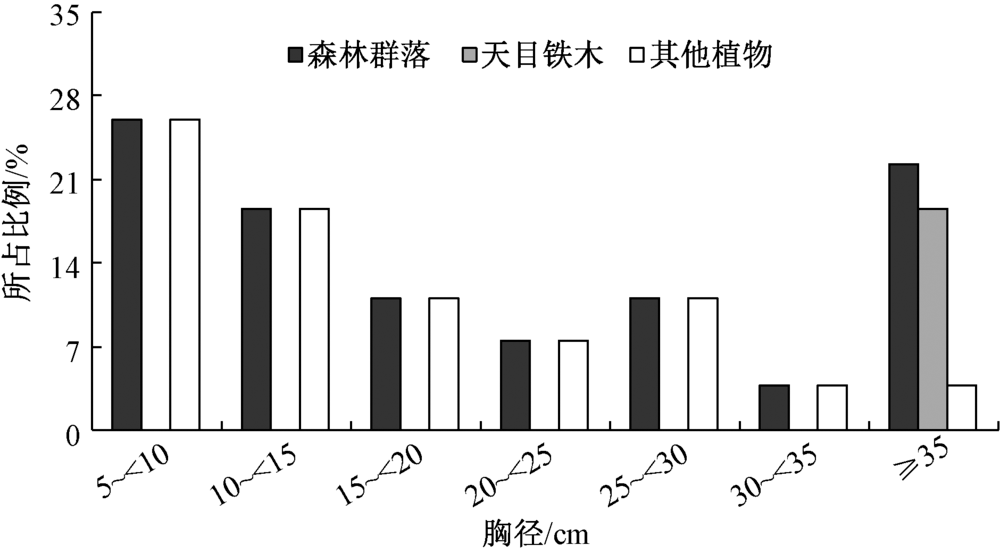 | 图3 乔木层树种的径阶分布 |
森林群落的垂直结构体现了不同树种的高度, 反映了不同植物在群落中的分层结构和生态位[17]。天目铁木群落是以天目铁木、马尾松为优势树种组成的常绿落叶针阔混交群落, 具有完整的乔木层、灌木层和草本层, 共有维管植物43科67属73种。
某一树种胸径大小可以反映其年龄水平[18], 可以用径阶结构代替时间顺序关系, 反映种群动态变化[19]。天目铁木群落中的天目铁木径阶结构呈现出明显的偏峰型, 所有个体均为老龄, 胸径均大于35 cm, 幼、中龄个体则为零, 表现为衰退型的种群特征。而马尾松、枫香等其他树种的径阶结构表现为“ 逆J-字型” , 即幼龄个体数量绝对优势, 表现为增长型的结构特征。在自然条件下, 种子向幼苗的转化是物种濒危的关键环节, 没有足够数量的幼苗, 种群就难以维持。种子产量低、品质差, 种子向幼苗的转化率低, 形成的幼苗数量少是频危植物存在的较普遍问题[20]。已有研究表明, 天目铁木的种子不饱满、品质低, 在野外环境下种子萌发率和成苗率均极低[8]。在调查中发现, 研究区域内天目铁木有一定数量的幼苗, 是人工繁育苗木的回归种植; 另外天目铁木种群位于公路边、山麓农舍旁的茶园、竹林中, 群落周围为栽培的高节竹林及稀疏的马尾松, 人为干扰强度高。
针对天目铁木的濒危现状, 该种群的保护和恢复策略主要有以下两点: 一是从个体自身因素考虑, 在加强对原生境及植株的保护与监测基础上, 加大科研力度, 开展天目铁木生殖生态学研究, 如种子萌发率、成苗率和高效繁育技术等, 提高实生苗的数量, 以扩大其种群分布范围; 人工种植, 引种栽培并加强推广, 扩大其生态位。二是从所处生境考虑, 加强对周边农民的宣传教育, 提高保护意识; 适度剔除竞争植物(清除树下及周围的竹子等)基础上, 采取封禁措施, 以恢复其自然更新环境, 保障天目铁木幼苗的生存空间。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|