


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Pb2+、Zn2+、Cd2+胁迫对盐地碱蓬种子萌发的影响
引用本文
邱振鲁, 吴杨艺璇, 孙桢, 于学文, 庄杰隆. Pb2+、Zn2+、Cd2+胁迫对盐地碱蓬种子萌发的影响 [J]. 浙江农业科学, 2018,59(12):2265-2269

doi: 10.16178/j.issn.0528-9017.20181238
Permissions
Copyright©2018, 《浙江农业科学》编辑部
《浙江农业科学》编辑部 所有
Pb2+、Zn2+、Cd2+胁迫对盐地碱蓬种子萌发的影响
作者简介:邱振鲁(1985—),男,山东济南人,讲师,硕士,研究方向为环境生物化学,E-mail:459139428@qq.com。
摘要
为研究Zn2+、Pb2+、Cd2+三种重金属离子单一胁迫对盐地碱蓬种子萌发的影响,以发芽势、发芽率、发芽指数、根长、芽长和根系活力为指标测定重金属离子对盐地碱蓬种子发芽的影响。结果表明,Pb2+和Cd2+浓度的升高对盐地碱蓬种子的发芽率和发芽势均有明显抑制作用,但Zn2+浓度的升高仅对盐地碱蓬的发芽率有明显抑制作用。随着3种重金属离子浓度的升高,发芽指数、根长和芽长均呈不同程度下降趋势,但Pb2+在低浓度下对根长的抑制程度不明显。Pb2+胁迫和Cd2+胁迫下的盐地碱蓬种子根系活力均随胁迫浓度增加呈整体下降趋势;低浓度Zn2+胁迫对盐地碱蓬种子的根系活力反而有促进作用,从中高浓度开始对盐地碱蓬种子的根系活力产生明显抑制作用。该研究结果可以为重金属污染区域土壤的盐地碱蓬种植提供一定的参考依据。
关键词:
发芽势; 发芽率; 根系活力; 盐地碱蓬; 重金属
中图分类号:S156.4+9
文献标志码:A
文章编号:0528-9017(2018)12-2265-05
盐地碱蓬(Suaeda salsa), 藜科碱蓬属植物[1], 一年生草本, 多分布在黄河三角洲海滨及荒漠低处的盐碱地上[2], 为辽宁、山东、江苏一带平原海岸淤滩上的优势物种[3]。研究表明, 种植盐地碱蓬能大大降低土壤的含盐量[4], 并且作为盐碱地指示生物, 对其进行研究有助于促进植物耐碱胁迫机理和应用的研究发展[5]。
近年来, 重金属污染日益严重, 很多盐碱地已经不是单纯的盐碱污染, 而是盐碱和多种重金属的复合污染, 所以, 研究重金属对植物生长发育的影响十分必要。重金属通过矿采、化工、化石燃料的燃烧、各类污水的排放、农药以及化肥的施用、大气沉降等途径持续地进入生物圈, 在环境中具有潜在危害, 极易被生物富集, 并在土壤中累积, 对种子的萌发过程产生不良影响[6]。目前关于盐地碱蓬种子萌发的研究, 仅限于盐碱胁迫下种子萌发指标与幼苗生长指标的研究[7], 以及盐碱与营养元素互作对上述指标影响的研究[8]和外源性物质浸种对盐胁迫下盐地碱蓬种子萌发指标的影响[9]。重金属胁迫对盐地碱蓬种子发芽各指标的影响未见文献报道。本论文研究了不同浓度Zn2+、Pb2+、Cd2+胁迫对盐地碱蓬种子萌发的影响, 以期为盐地碱蓬在重金属污染环境下的种植以及盐生植物资源的开发利用提供参考。
1 材料与方法
1.1 材料
盐地碱蓬种子(上海源叶生物科技有限公司提供), Pb(NO3)2、ZnSO4、CdCl2均为分析纯。
1.2 方法
1.2.1 种子的选取与预处理
盐地碱蓬种子具有二型性, 本实验采用的是黑色的籽粒。用镊子挑选大小相近、无破损、发黑发亮的种子, 用2% NaClO浸泡30 min, 除去种子表面的杂菌。再用蒸馏水反复冲洗, 洗净残留在表面的NaClO, 风干备用。
1.2.2 处理设计
分别配置浓度为0.1、0.2、0.4、0.6、0.8、1.6、3.2 g· L-1的ZnSO4工作液[10], 0.05、0.1、0.2、0.3、0.5、0.8、1.2 g· L-1的Pb(NO3)2工作液[11]和0.01、0.05、0.1、0.2、0.3、0.4、0.5 g· L-1的CdCl2工作液[12]。
向经高压蒸汽灭菌的培养皿中放入滤纸, 分别加入上述不同浓度和种类的工作液, 排出滤纸内气泡并均匀放入处理好的种子50粒, 盖好培养皿, 于25 ℃恒温培养箱中培养[13]。每组处理做3个重复。
1.2.3 指标测定
测定指标包括发芽率、发芽势、发芽指数、根长和芽长及根系活力[14, 15]。以胚根突破种皮1 mm视为发芽, 发芽指数计算到14 d。发芽14 d时, 测定根长和芽长, 以种子开始变色且翘起的位置至最长叶叶尖的长度为芽长, 剩余的位置至最长根根尖的长度为根长, 每皿选取长度接近的10株进行测量, 取平均值[16]。
根系活力测定采用TTC还原法, 制作TTC标准曲线, 以空白实验样品液为参比, 在485 nm波长下测定剪取的根尖样品处理液的吸光度, 根据吸光度查标准曲线, 即求出TTC还原量, 按照以下公式即可求出根系活力[17]:
TTC还原强度(mg· g-1· h-1)=m/(m0· t)
式中:m为从标准曲线查得的TTC还原量(mg); m0为根尖样品质量(g); t为反应时间(h)。
2 结果与分析
2.1 对盐地碱蓬发芽率及发芽势的影响
2.1.1 Pb2+
不同浓度Pb2+对种子发芽率和发芽势的影响如图1所示, 不同浓度处理组之间发芽势差异均显著, 其中, Pb2+浓度为0.05和0.10 g· L-1时, 发芽势与对照组相比差异极显著, 可知0.05 g· L-1Pb(NO3)2胁迫就可以对盐地碱蓬的发芽势产生抑制作用。将其处理分为低浓度组(0.05、0.1 g· L-1)、中浓度组(0.2、0.3、0.5 g· L-1)和高浓度组(0.8、1.2 g· L-1), 数据分析表明, 组间差异极显著, 组内差异显著。不同浓度处理组之间的发芽率差异显著性分析结果表明, 与对照相比, 低浓度组内Pb2+对发芽率影响显著, 中浓度组内及高浓度组内差异不显著, 但组间差异显著。
 | 图1 Pb2+、Zn2+、Cd2+胁迫对盐地碱蓬发芽势和发芽率的影响 |
不同浓度Pb2+胁迫下, 发芽势的波动幅度大于发芽率的波动幅度, 说明Pb2+对发芽势影响更大, 即对种子萌发前期的影响更为明显。
2.1.2 Zn2+
从发芽势来看, 对照组与低浓度处理(0.1 g· L-1)和中等浓度处理组内(0.2、0.4、0.6 g· L-1)差异不显著, 但组间差异极显著。从发芽率来看, Zn2+浓度为0.1 g· L-1时, 发芽率极显著低于对照组; 另外, 在相邻浓度的处理中, 即0.2 g· L-1和0.4 g· L-1、0.6 g· L-1和0.8 g· L-1以及0.8 g· L-1和1.6 g· L-1之间, 发芽率表现出极显著差异, 其他相邻处理组之间差异均显著, 其中0.1 g· L-1和0.2 g· L-1处理组的发芽率差异不显著。ZnSO4在低浓度下对发芽势具有抑制作用, 但中高浓度相比中低浓度呈上下波动状态。随ZnSO4浓度的升高发芽率呈降低趋势。比较发芽势和发芽率变化趋势可知, 低浓度Zn2+对发芽势影响更加明显, 而高浓度Zn2+对发芽率影响更明显。
2.1.3 Cd2+
0.05 g· L-1处理组与0.01 g· L-1处理组发芽势差异显著, 而0.01 g· L-1处理组与对照组差异不显著, 说明CdCl2从0.05 g· L-1开始对盐地碱蓬的发芽势产生抑制作用。CdCl2超过0.05 g· L-1后, 每个处理组都和相邻低浓度处理组之间差异显著, 其中, 0.2 g· L-1的处理组与0.3 g· L-1的处理组之间发芽势差异极显著。发芽率数据分析表明, CdCl2浓度为0.01 g· L-1时, 发芽率极显著低于对照组, 表明从该浓度起, Cd2+就对发芽率产生了明显的影响。从0.2 g· L-1到0.4 g· L-1, 每个处理组的发芽率均与前一个处理组差异极显著, 其余处理组之间发芽率无显著性差异。不同浓度Cd2+胁迫下:发芽势整体呈波动下降趋势, 其中, 0.05 g· L-1处理组与前一组比较, 作用最为明显; 发芽率呈持续下降趋势。
2.2 对发芽指数的影响
Pb2+、Zn2+、Cd2+胁迫下盐地碱蓬发芽指数的变化趋势如图2所示, Pb(NO3)2胁迫下盐地碱蓬种子发芽指数持续下降, 说明随着Pb(NO3)2浓度的升高, 能使种子在失去发芽力之前持续劣变, 种子活力持续下降; 随ZnSO4浓度的升高, 发芽指数整体呈下降趋势, 在0.1~0.6 g· L-1时, 下降趋势更为明显。随着CdCl2浓度的升高, 发芽指数也呈整体下降趋势, 在0.1~0.5 g· L-1时, 下降趋势最明显。
 | 图2 Pb2+、Zn2+、Cd2+胁迫对盐地碱蓬发芽指数的影响 |
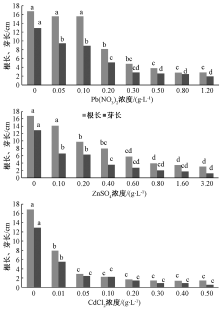
2.3 对根长和芽长的影响
2.3.1 Pb2+
由图3可知, 从0.2 g· L-1开始, Pb2+对根长有显著抑制作用; Pb2+浓度高于0.5 g· L-1时, 根长显著小于0.2 g· L-1的处理组。Pb2+浓度为0.05 g· L-1时, 芽长显著小于对照组, 说明0.05 g· L-1的Pb2+就开始抑制盐地碱蓬芽的生长; 随着Pb2+浓度的升高, 中低浓度处理组芽长差异显著, 而高浓度组之间差异不显著。Pb2+胁迫对根长和芽长的影响趋势大致相同, 均表现为随浓度增加而下降, 尤其是在中等浓度变化范围内, 下降趋势更为明显。
 | 图3 Pb2+、Zn2+、Cd2+胁迫对盐地碱蓬根长和芽长的影响 |
2.3.2 Zn2+
Zn2+浓度为0.2 g· L-1的处理组盐地碱蓬根长显著低于0.1 g· L-1和对照组, 说明Zn2+浓度达到0.2 g· L-1时, 开始对根尖细胞分裂产生明显的抑制作用; Zn2+浓度高于0.8 g· L-1的处理组之间根长均无显著差异。0.2 g· L-1处理组与0.6 g· L-1处理组之间差异显著, 且均与0.4 g· L-1处理组之间差异不显著。随胁迫Zn2+浓度的增加, 盐地碱蓬根长和芽长均呈现明显的下降趋势, 根长的下降趋势随Zn2+浓度增加比较稳定, 而在较低的浓度下即可强烈抑制盐地碱蓬芽的细胞分裂。
2.3.3 Cd2+
Cd2+胁迫下, 盐地碱蓬根长和芽长的变化趋势一致, 0.01 g· L-1处理组与对照组及0.05 g· L-1处理组之间差异显著, 说明低浓度Cd2+处理已经对根尖和芽的细胞分裂产生了显著的抑制作用; 而Cd2+高于0.05 g· L-1的处理组之间根长、芽长差异不显著, 这可能是低浓度Cd2+的抑制效应几乎已经达到极限, 从而使高浓度抑制增强的作用不明显。
2.4 对盐地碱蓬根系活力的影响
2.4.1 Pb2+
盐地碱蓬的根系活力随Pb2+浓度的增加而呈现持续下降趋势, Pb2+浓度超过0.3 g· L-1时, 根系活力下降趋势更大, 说明实验中设置的高浓度处理组对根系中各种脱氢酶的抑制作用更加明显(图4)。
 | 图4 Pb2+、Zn2+、Cd2+胁迫对盐地碱蓬根系活力的影响 |
2.4.2 Zn2+
与Pb2+胁迫的结果不同, 低浓度Zn2+胁迫可以提高盐地碱蓬根系活力, 而高浓度Zn2+胁迫强烈抑制根系活力, 推测这与锌是植物生长的必要元素有关。研究表明, 锌是甘油醛-3-磷酸脱氢酶的活化剂, 低浓度Zn2+胁迫能够促进根系活力, 而高浓度Zn2+的抑制作用可能与重金属引起胞内蛋白变性或对某些根系活力指标中脱氢酶的抑制作用有关。
2.4.3 Cd2+
盐地碱蓬的根系活力随Cd2+浓度增加呈现持续下降趋势, Cd2+浓度为0.05~0.3 g· L-1时, 随浓度增加, 根系还原酶活力下降最为明显。
3 讨论
Pb2+对盐地碱蓬发芽率和发芽势均呈现始终抑制的作用, Pb2+浓度为0.05 g· L-1时便与对照组产生了显著差异, 这说明Pb2+在较低的浓度下, 可以抑制种子萌发时相关酶的活力或者使细胞内蛋白或核酸变性。Zn2+对盐地碱蓬发芽势表现出低浓度促进而高浓度抑制的结果, 可能是Zn2+作为植物的必须元素, 低浓度时能够激活种子萌发时某些酶的活性, 高浓度时导致胞内蛋白变性和对种子萌发过程中酶有非竞争性抑制作用。Zn2+对发芽率的影响没有出现低浓度促进的现象, 猜测其原因可能是由于种子已经在Zn2+胁迫处理14 d, 其细胞内的Zn2+积累较多。低浓度Cd2+胁迫即可以显著抑制盐地碱蓬发芽率和发芽势, 随Cd2+浓度增大, 盐地碱蓬发芽率和发芽势受抑制程度均增加, 推测与重金属导致核酸、蛋白变性或者抑制酶的活性有关。
Pb2+、Zn2+、Cd2+对盐地碱蓬发芽指数的影响与对发芽率、发芽势的影响几乎一致, 原因也可以认为同上述分析。而对根长和芽长的影响, 也与这3种重金属对发芽率的影响几乎一致。根的伸长是根尖分生区细胞分裂和伸长区细胞伸长的结果, 芽的增长是芽细胞分裂伸长的结果, 重金属进入细胞, 使核酸或蛋白变性, 或者抑制有丝分裂过程中纺锤体的形成从而抑制细胞的分裂, 或者通过抑制细胞骨架蛋白的形成影响细胞的伸长, 可能是导致根长、芽长随重金属浓度增加而下降的原因。
除低浓度Zn2+对盐地碱蓬根系活力有略微的促进作用外, Pb2+、Cd2+均表现为随浓度的升高对根系活力的抑制作用增强, Zn2+超过0.4 g· L-1后, 开始对根系活力呈现抑制作用。低浓度Zn2+使根系活力增强的原因可能是因为其作为甘油醛-3-磷酸脱氢酶和乙醇脱氢酶的组成成分, 它们在细胞内的合成量增加从而使酶活性增强, 也可能是Zn2+作为脱氢酶的激活剂使酶活性增强。
4 小结
本文使用平皿培养法, 研究3种重金属Pb2+、Zn2+和Cd2+单一胁迫对盐地碱蓬种子萌发的影响, 结果表明, 只有Zn2+在低浓度下对盐地碱蓬种子的发芽势、发芽指数和根系活力有促进作用。Pb2+、Cd2+对种子萌发各指标的影响均表现为随浓度的升高抑制作用逐渐增强, Zn2+在高浓度下对种子萌发各指标的抑制作用也随浓度增加而增强。
The authors have declared that no competing interests exist.
参考文献:
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|