

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
烟草 NtSYP121基因克隆、表达载体构建及表达分析
[熊雪 , 殷双琴, 陈欣, 王丽红, 宋文强, 李立芹
, 殷双琴, 陈欣, 王丽红, 宋文强, 李立芹* ]
, 殷双琴, 陈欣, 王丽红, 宋文强, 李立芹]
|
|


作者简介:熊 雪(1996—),女,四川眉山人,本科生,从事植物分子生物学研究工作,E-mail:742193922@qq.com。
采用同源克隆的方法对烟草品种K326中的一个 SYP121基因进行了研究,该基因全长903 bp,蛋白编码300个氨基酸。同源性分析结果显示,该基因所编码的蛋白与渐窄叶烟草SYP121-like、绒毛状烟草SYP121-like等的亲缘关系最近,故命名为 NtSYP121。组织表达分析发现,该基因在成熟期的根、茎、叶、花中均有表达,在根中的表达量最高。非生物逆境胁迫实验表明,该基因对低钾、高盐、干旱、ABA均有响应表达,并成功构建了 NtSYP121 -pBI121过表达载体。研究结果为解析 NtSYP121在响应逆境胁迫的功能奠定一定理论基础。
植物本身无法主动逃避不利环境, 为适应各种外来以及内在胁迫, 植物细胞不断调整并最终形成了特有的内膜系统和膜泡运输机制以完成细胞内和细胞间的物质交流[1]。植物 SNARE (soluble N-ethylmaleimide-sensitive fusion protein attachment protein receptor)类蛋白是一类具有抗生物和非生物逆境功能的蛋白。目前已经发现许多植物SNARE蛋白可通过调节离子通道、控制蛋白质密度、直接与受体作用等方式来调节离子运输, 从而对刺激性反应作出应答[2]。SNAREs蛋白是可溶性NSF附着蛋白受体, 是介导钙离子依赖的囊泡膜与突触膜融合的核心分子复合体[3]。SNARE蛋白的分类方法有两种:根据膜融合过程中SNARE蛋白所处位置, 可分为v-SNAREs和t-SNAREs[4]; 而根据SNAREs模体(motif)氨基酸的不同, 又可将SNAREs分两类:Q(谷氨酰胺)和R(精氨酸), Q-SNARE包括Syntaxin和SNAP25两个亚家族; R-SNARE包括synaptobrivin/vamp亚家族。拟南芥基因组编码24个syntaxin型Q-SNARE, 3个SNAP25型Q-SNARE, 14个VAMP型R-SNARE。Syntaxin型Q-SNARE又可细分为8个亚家族(Syp1-Syp8)[2]。
SYP121是亚家族蛋白Syntaxin中的一员。在拟南芥中, SYP121调控K+通道的亚基KC1, 并与AKT1形成三元复合物SYP121-KCl-AKT1共同参与植物的钾营养[5]。Grefen[6]发现SYP121和KC1互作是与一个FxRF模体相关联的, 而这个FxRF模体独特地位于SNARE序列的前12个残基。Geelen等[7]发现, SYP121介导了高尔基复合体和质膜之间的囊泡运输。也有研究表明, SYP121的功能失活能改变KATI钾通道的定位[8]。此外, AtSYP121还可以提高植物的抗病性。例如, Kwon等[9]研究发现, AtSYP121能与SNAP33和VAMP721/VAMP722形成复合体, 共同调控病原体侵染时的细胞分泌。Maekawa等[10]发现SYP121参与拟南芥抵抗白粉病生理过程。Kiyono等[11]的研究结果表明过量表达SYP121和重金属转运体MerC融合蛋白能增加拟南芥中镉积累和耐受性。在烟草中, Leyman等[12]研究发现SYP121在保卫细胞中调控K+和Cl-通道, 推测在脱落酸信号中起作用。
综上所述, Syntaxin家族蛋白对植物的抗逆性起重要作用, 但是在烟草中的相关研究却比较少。因此, 本研究利用K326烟草克隆了NtSYP121基因对并对其序列进行生物信息学分析, 同时, 运用Real-time PCR分析了其在根、茎、叶、花及低钾、高盐、干旱和ABA处理下基因的表达水平, 并成功构建NtSYP121-pBI121过表达载体, 这对于NtSYP121响应逆境胁迫的功能研究具有重要意义。
实验所用材料是K326烟草, 由四川农业大学植物生理与生物技术系提供。首先进行漂浮育苗, 待其生长到十字期, 将其移栽到花盆中, 每盆种植一株。对烟株正常生长的根、茎、叶和花进行取样, 然后在液氮中速冻, 放置-80 ℃冰箱, 以供后续试验使用。
将烟草品种K326种子表面消毒后布种于MS培养基上, 培养15 d后挑取长势一致的幼苗分别进行ABA、PEG、高盐和低钾处理, 每个处理设有3个重复。其中, ABA、PEG及高盐处理培养基是在MS培养基的基础上分别加入ABA(1 μ mol· L-1)、PEG6000(5%)、NaCl(200 mmol· L-1)处理; 低钾处理时将待处理材料转移到低钾培养基上, 低钾培养基是将MS培养基中的KNO3去掉, 并且将KH2PO4替换为NH4H2PO4, 其余成分相同, 经测定钾离子浓度为10 μ mol· L-1。所有处理在0、3、6、12、24 h后进行整株取样。
通用型DNA纯化回收试剂盒购自天根公司; DH5α 大肠埃希菌菌株购于Vazyme Biotech公司; Trizol试剂、cDNA合成试剂盒、载体PMD19-T、高保真Pfu酶、PrimeScript RT reagent Kit、SYBR Green Master mix、限制性内切酶、DNA Ligation Kit 2.0等试剂购自TaKaRa生物公司, 引物合成与测序由成都梓熙擎科公司完成。
1.2.1 克隆目的基因
取幼嫩的烟草叶片于液氮中速冻1 min后用Trizol试剂法提取其总RNA, 再根据cDNA合成试剂盒的操作要求, 将其反转录成cDNA。反转录体系为10 μ L, 在EP管中依次加入0.5 μ L OligodT, 5 μ L RNA, 0.5 μ L无菌水, 轻微摇匀后, 在PCR仪上65 ℃变性5 min, 再在冰上急冷2 min; 然后再依次加入2 μ L 5× Reaction Buffer, 0.5 μ L RNase Inhbitor, 1 μ L 10 mmol· L-1 dNTP, 0.5 μ L Reverse Transcripase, 在PCR仪上42 ℃反应60 min, 72 ℃延伸10 min后终止反应, 取出1 μ L稀释10倍备用, 其余-20 ℃保存。
参考NCBI GenBank收录的普通烟草NtSYP121序列(登录号:NM_001325000), 采用同源克隆的方法用DNAMAN设计引物NtSYP121-F和NtSYP121-R(表1), 再以其稀释过的cDNA为模板进行PCR扩增。反应体系为10 μ L, 分别加入5 μ L 2× Perime STAR Mixture, 0.2 μ L 引物F (10 μ mol· L-1), 0.2 μ L 引物R (10 μ mol· L-1), 1 μ L cDNA, 3.6 μ L ddH2O。反应程序为:95 ℃预变性5 min, 1个循环; 95 ℃变性30 s, 57 ℃退火30 s, 72 ℃延伸2.5 min, 35个循环; 72 ℃ 8 min, 1个循环; 4 ℃永久保存。
1.2.2 PCR产物的回收、克隆与测序
PCR产物经1.2%琼脂糖凝胶电泳检测后, 将目的条带按照TIANGEN Universal DNA Purification Kit的说明操作进行纯化回收, 将目的片段与pMD19-T载体16 ℃连接过夜, 连接产物转化大肠埃希菌DH5α 感受态, 随后在涂有含氨苄青霉素(Ampicillin)的LB平板上进行筛选, 采用菌落PCR法检测阳性克隆, 然后送到成都梓熙擎科公司进行测序。
1.2.3 目的基因的生物信息学分析
使用ExPASyProtParam tool(http://web.expasy.org/protparam/)对NtSYP121基因编码的蛋白质进行理化性质分析; 利用 DNAMAN 软件分析其编码蛋白的疏水性, 进行蛋白序列比对, 并构建系统进化树; 运用(http://www.ncbi.nlm.nih.gov/struc-ture/cdd/wrpsb.cgi)进行保守功能域分析; 采用PSORT (http://psort.hgc.jp/form.html)预测NtSYP121的亚细胞定位; 采用IBCP(http://npsa-pbil.ibcp.fr)的在线工具SOPMA预测二级结构; 使用SWISS-MODEL(http://swissmodel.expasy.org)在线预测三级结构。
1.2.4 NtSYP121-pBI121过表达载体构建
采用全长含有酶切位点引物NtSYP121-pBI121-F和NtSYP121-pBI121-R扩增目的基因(表1), 扩增结束后, 将PCR产物和pBI121载体分别进行双酶切, 酶切体系10 μ L:10× T Buffer 0.5 μ L, BSA 1 μ L, BamHⅠ 1 μ L; SmaⅠ 1 μ L; PCR产物/pBI121载体 4 μ L; ddH2O 2.5 μ L。酶切后回收目的片段和pBI121载体, 于16 ℃连接过夜, 转化大肠埃希菌DH5α 感受态, 随后在含有卡那霉素的LB平板上进行筛选, 菌落PCR检测筛选阳性克隆后, 摇菌提取质粒, 双酶切后送至成都梓熙擎科公司进行测序。
1.2.5 基因的表达模式分析
根据目的基因的序列, 使用DNAMAN软件设计其荧光定量PCR的引物NtSYP121qF和NtSYP121qR(表1)。提取烟草总RNA后进行反转录, 获得cDNA, 再以cDNA为模板进行荧光定量PCR反应, 反应体系为25 μ L, 依次加入:SYBRⅡ 12.5 μ L, 引物F、R各1 μ L, cDNA 1.5 μ L, ddH2O 9 μ L。反应程序为95 ℃预变性3 min, 1个循环; 95 ℃变性5 s, 60 ℃延伸45 s, 40个循环。所有样品都设置3个重复来扩增目的基因, 以烟草18S基因设计引物作为实验的内标定量, 扩增目的基因。扩增结束后, 按2-△ △ Ct法计算相对表达量。
| 表1 各基因的引物序列 |
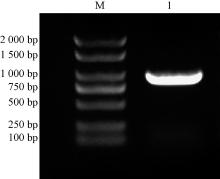
以烟草叶片总RNA反转录合成的cDNA为模板进行PCR扩增, 电泳结果显示在1 000 bp左右的位置上有清晰的条带(图1)。采用切胶回收的方法对该条带进行纯化后, 与pMD19-T载体16 ℃连接过夜, 随后对获得阳性克隆随机挑选3个进行测序。测序结果显示:目的片段大小为903 bp。
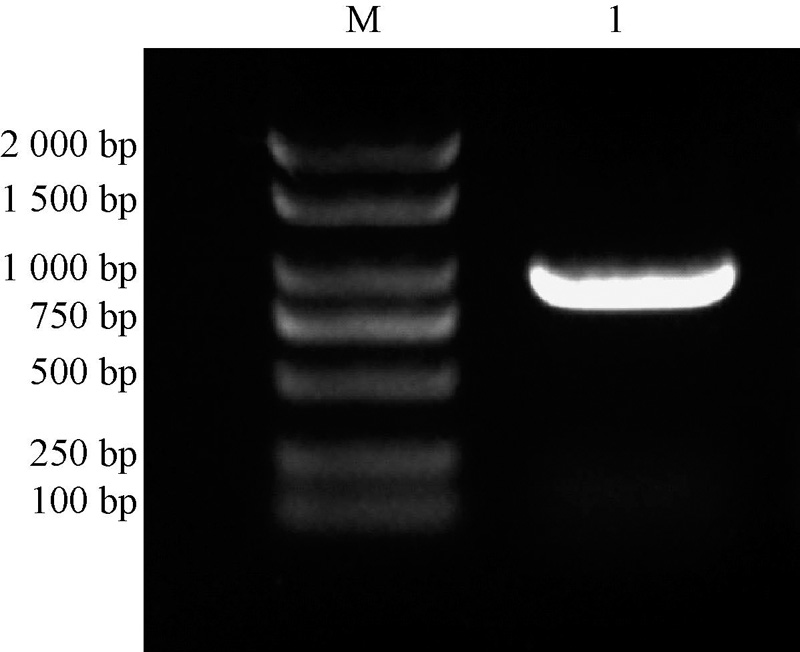 | 图1 NtSYP121的克隆 M为Marker 2 000 bp; 1为NtSYP121 |
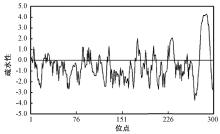
使用ExPASyProtParam tool(http://web.expasy.org/protparam/)对NtSYP121基因编码的蛋白质进行了理化性质分析, 结果表明, 该蛋白质的分子量为34 ku, 理论等电点pI为7.76; 带正电荷的氨基酸残基(精氨酸+赖氨酸)总数是43, 带负电荷的氨基酸残基(天冬氨酸和谷氨酸)总数是42, 该蛋白的不稳定系数是40.07, 由此说明该蛋白可能是一个不稳定蛋白。该蛋白脂肪系数为85.13, 总的平均疏水性为-0.551, 用DNAMAN软件进行的疏水性分析结果显示(图2), 预测该蛋白属于亲水性蛋白。
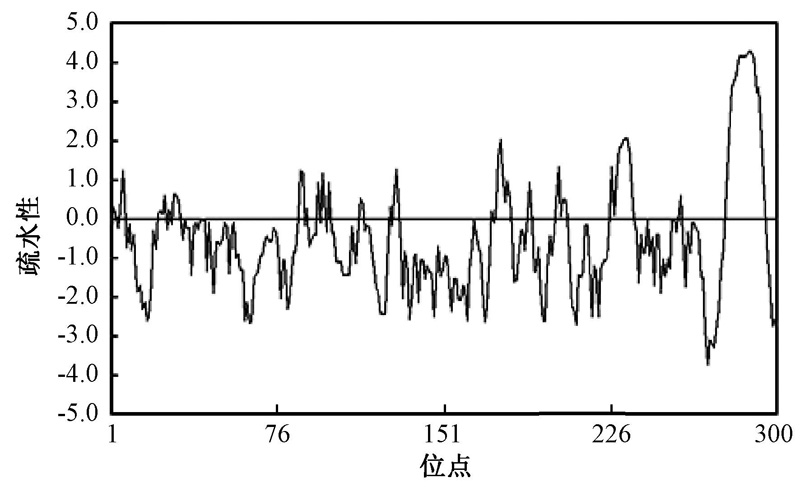 | 图2 NtSYP121疏水区的预测 |
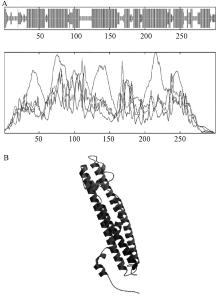
使用IBCP(http://npsa-pbil.ibcp.fr)的在线工具SOPMA, 对NtSYP121的二级结构进行预测, 结果(图3中A)表明:α -螺旋约占65.00%, 无规则卷曲约占20.00%, 延伸链占8.33%, β -转角占6.67%, 由此可见, 该蛋白二级结构的最大元件为α -螺旋。使用SWISS-MODEL对NtSYP121的三级结构进行了预测, 结果如图3中B所示, 并将结果与二级结构预测及跨膜结构预测比照, 结果较为统一, 所得三级结构预测结果较为可信。利用 Swiss-PdbViewer分析同源建模结果, 表明其立体三维结果比较稳定, 所以其同源建模的结果是相对可靠的。
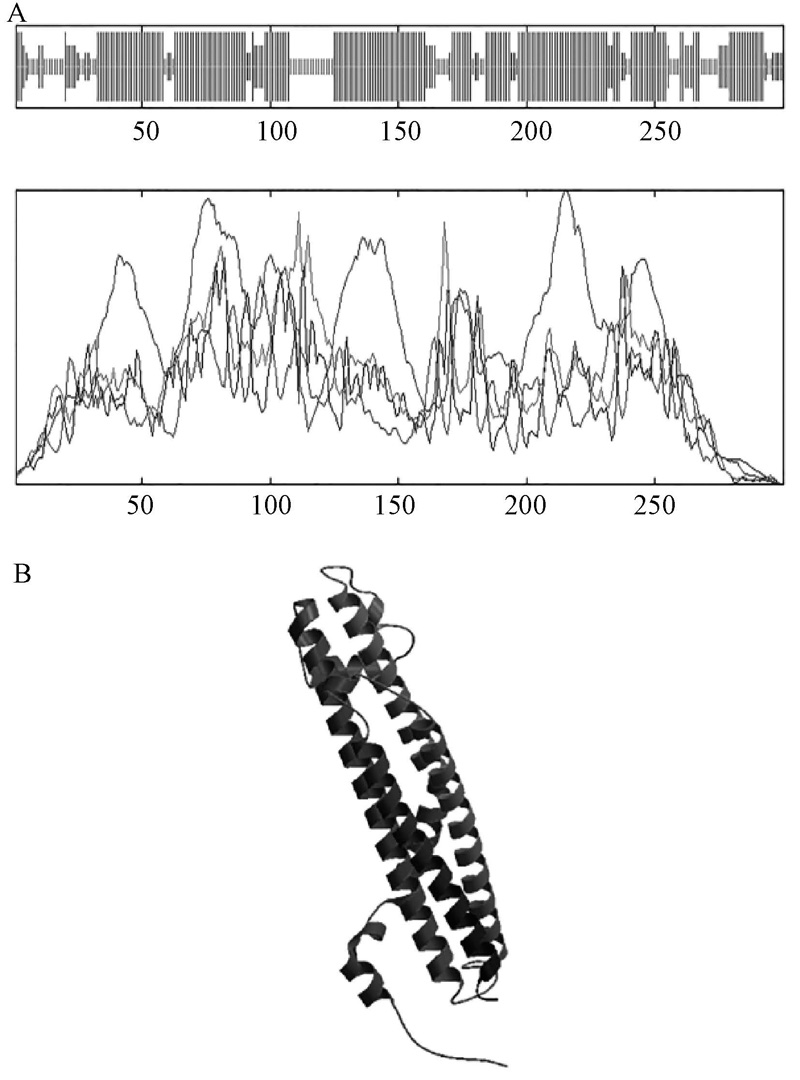 | 图3 NtSYP121的二级结构及三级结构预测 A, 二级结构预测; B, 三级结构预测 |
为了探明NtSYP121与其他植物基因的同源性, 把NtSYP121编码的氨基酸序列在NCBI 数据库中进行了BLAST比对, 然后在DNAMAN进行多序列比对, 结果(图4中A)显示, NtSYP121与各植物中的SYP121蛋白一级结构的氨基酸序列是高度保守的。将NtSYP121氨基酸序列与其他同源性比较近的植物SYP121序列进行聚类分析, 结果表明, NtSYP121与渐窄叶烟草SYP121-like、绒毛状烟草SYP121-like、番茄SYP121-like等具有较高的氨基酸序列同源性, 分别为99%、98%、94%(图4中B)。
利用NCBI数据库的CDD软件对目的基因所编码蛋白的功能保守域进行了分析, 结果(图5)表明, 该蛋白具有SNARE-Syntaxin家族典型的N端结构, 且属于t-SNARE蛋白, 说明其是多功能蛋白家族SNARE的亚家族蛋白Syntaxin的一员。使用PSORT(http://psort.hgc.jp/form.html)在线预测亚细胞定位, 根据预测的结果得知, 在质膜上的可能性为0.440, 在线粒体内膜上的可能性为0.100, 在高尔基体上的可能性为0.100, 在内质网膜上的可能性为0.850。根据以上结果预测该蛋白质定位在内质网膜上。
 | 图4 NtSYP121同源性分析 A:NtSYP121蛋白多重序列比对; B:NtSYP121蛋白系统进化树 |
 | 图5 目的基因编码蛋白的功能保守域分析 |
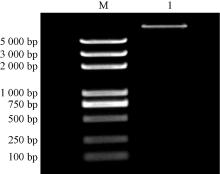
将PCR扩增产物和pBI121载体分别进行双酶切, 回收后连接并转化大肠埃希菌, 培养阳性菌落进行质粒提取, 然后进行双酶切, 电泳结果(表6)显示, 能够酶切出目的基因的条带, 表明NtSYP121-pBI121过表达载体构建成功。
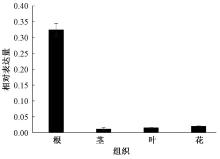
以普通烟草K326的根、茎、叶、花cDNA为模板进行Real-time PCR反应, 分析NtSYP121在各个部位的表达量, 结果(图7)表明, NtSYP121的表达量由高到低依次为根> 花> 叶> 茎, 其在根中的表达量是分别是花、叶、茎中表达量的15.0、20.2、26.4倍, 这说明NtSYP121基因主要在烟草植株的根中表达, 并且其表达存在组织特异性。
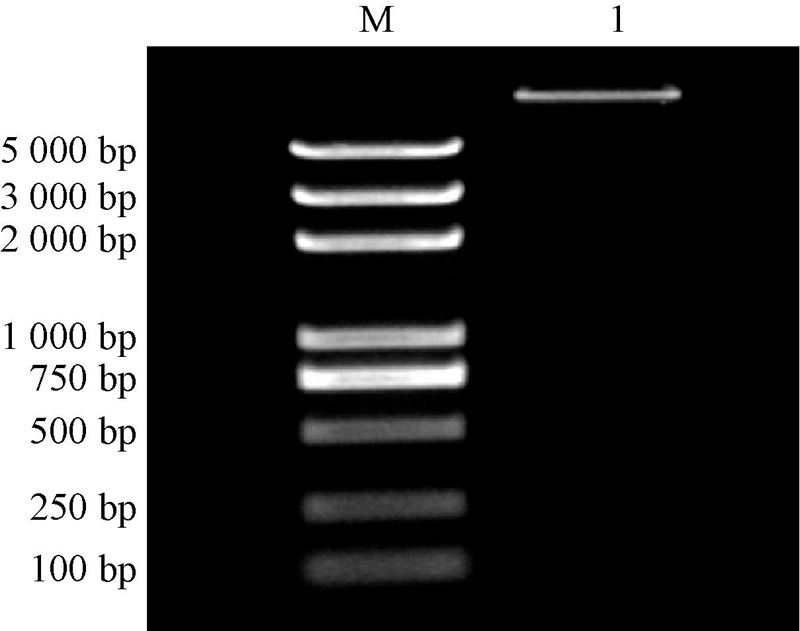 | 图6 NtSYP121-pBI121过表达载体构建 M, Marker 5 000 bp; 1, NtSYP121 |
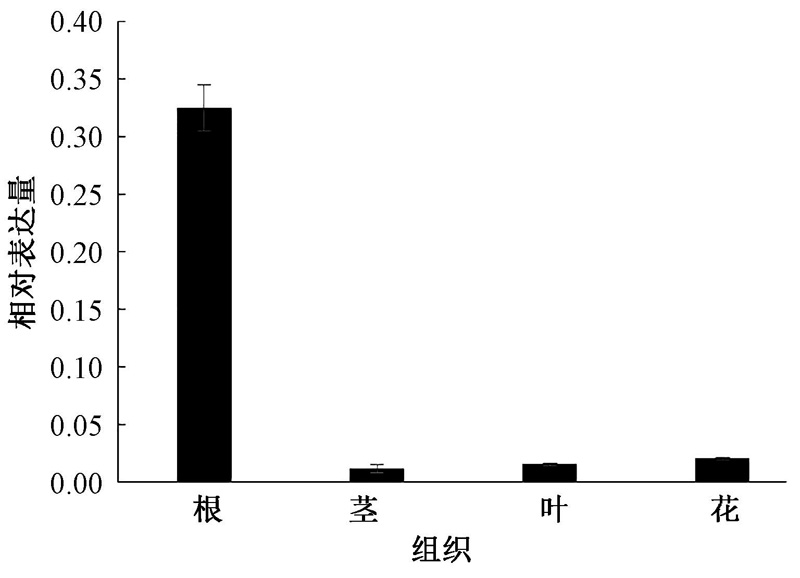 | 图7 NtSYP121在不同组织中的相对表达量分析 |
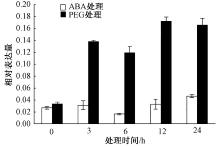
将烟草K326幼苗移苗至含有1 μ mol· L-1 ABA和5% PEG6000MS培养基上处理3、6、12、24 h, 然后进行取样提取RNA。采用Real-time PCR对NtSYP121在逆境胁迫下的表达模式进行分析。结果发现, 在ABA处理后, NtSYP121在3 h的表达量上调, 随后在6 h的表达量下降且低于对照(0 h), 在6 h和24 h表达量又逐渐上调, 其中在24 h时NtSYP121的表达量上调到最大值, 为对照(0 h)的1.72倍; 在PEG处理后, NtSYP121在3 h表达量明显增加, 其中在24 h时该基因表达量达到最大值, 为对照(0 h)的4.70倍。因此NtSYP121参与ABA和PEG胁迫生理过程(图8)。
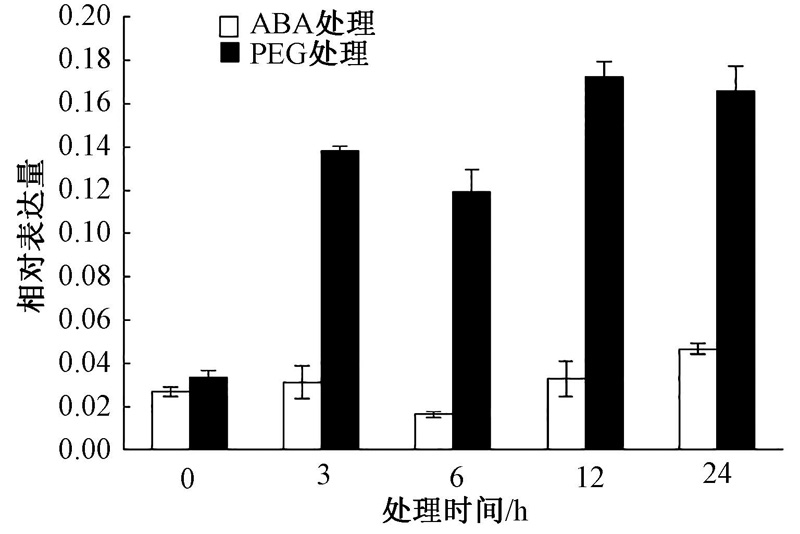 | 图8 ABA和PEG处理下NtSYP121的表达模式分析 |
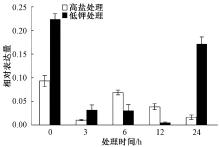
把生长15 d的烟草幼苗转移到高盐和低钾培养基上处理3、6、12和24 h, 然后进行取样提取RNA。采用Real-time PCR对NtSYP121在逆境胁迫下的表达模式进行分析, 结果发现, 在高盐处理后, NtSYP121在3 h的表达量下降, 仅为对照(0 h)的11%; 随后表达量上调, 6 h时表达量上调为对照的74%; 12 h及以后表达量又继续下降。在低钾处理3、6、12 h后, NtSYP121表达量都是下降的, 12 h时最低, 为对照(0 h)的2%; 随后24 h时表达量上调为对照的77%。上述结果表明高盐和低钾处理下NtSYP121表达量明显降低(图9)。
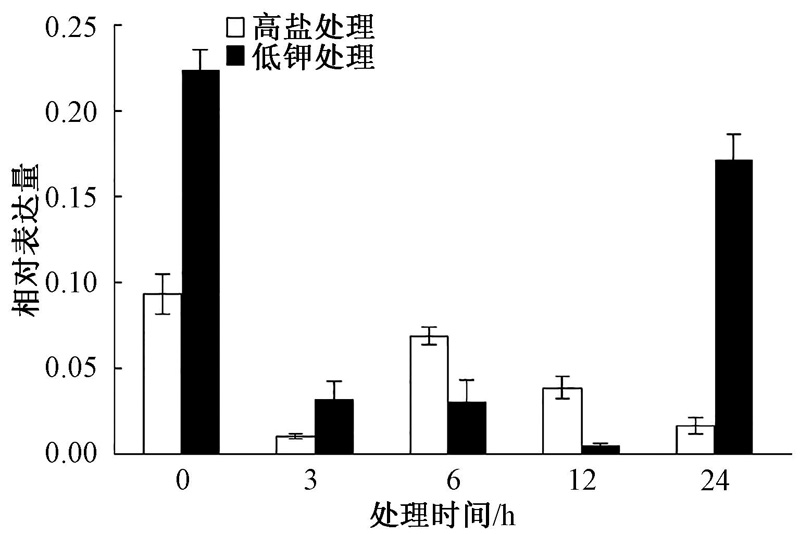 | 图9 高盐和低钾处理下NtSYP121的表达模式分析 |
本研究从普通烟草K326中克隆到一个Syntaxin家族成员NtSYP121的cDNA序列, 其ORF为903 bp, 编码300个氨基酸。编码蛋白的功能保守域分析, 该基因编码的蛋白具有SNARE-Syntaxin家族典型的结构。利用软件分析该蛋白的亚细胞定位, 表明该蛋白质定位在内质网膜上。而在Collins等[13]和Leyman等[14]对拟南芥和烟草的研究中, SYP121基因编码的蛋白定位在质膜上; 李文琦[15]对木薯MeSYP121蛋白进行亚细胞定位分析, 发现其定位到高尔基体中。今后将通过定位实验(如绿色荧光蛋白标记)来研究NtSYP121蛋白的定位, 这对于其功能的研究具有重要的意义。组织表达分析结果表明, NtSYP121基因的表达具有组织特异性, 在烟草K326的根中表达量最高, 而在茎、叶、花中表达量很低, 其中在茎中的表达量最低。Leyman等[14]在烟草Nicotiana的研究中也发现该基因所编码的蛋白在根中表达量最高, 而茎、叶、花中表达量较低, 本试验结果与前人研究结果一致, 因此推测NtSYP121主要在烟草根中发挥功能。
植物经历干旱胁迫时, ABA被普遍认为是一种干旱信号而传递干旱信息。逆境胁迫下, ABA通过信号转导通路, 诱导相关基因和蛋白表达, 从而提高植物对多种逆境胁迫的抗性[16, 17]。已有研究表明, ABA能提高植物对干旱胁迫的抗性[18, 19, 20]。在ABA(1 μ mol· L-1)处理后, NtSYP121基因表达量表现为在3 h上升, 在6 h时下降且低于对照, 随后该基因表达量又逐渐上升且高于对照, 最后在24 h时表达量最高; Real-time PCR分析结果表明:PEG处理后, NtSYP121的表达受到显著诱导, 其中在12 h时该基因表达量最高; 这与Leyman等[12]研究结果类似。在NaCl(200 mmol· L-1)和低钾胁迫处理后, Real-time PCR结果表明NtSYP121基因的表达量与对照相比, 呈下降趋势, 而在Leyman等[14]在烟草Nicotiana的研究中, 使用Northern blot方法检测该基因的表达量在NaCl(300 mmol· L-1)处理1 h升高, 使用Western blot方法测得该蛋白的表达量在6 h明显升高, 而后逐渐降低, 在24 h时表达量与对照没有区别。导致试验结果不同的原因可能是植物的生长时期, 盐处理浓度和检测方法的不同。要明确NtSYP121参与的信号转导途径还需要进一步的试验验证。已成功构建NtCBL1基因的过量表达载体, 后续工作可通过转基因技术研究该基因的功能。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|