
{kind=link}
8个玉簪品种幼叶的营养成分测定及品质分析
[马广莹1  , 史小华
, 史小华1 , 邹清成1 , 田丹青1 , 朱开元1 , 詹菁2 , 周江华1 ]
, 史小华|
|


作者简介:马广莹(1982—),男,博士,从事园林植物栽培育种研究工作,E-mail:mgyflower@163.com。
近年来,大量玉簪品种引进中国,为了挖掘其经济价值,拓宽应用领域,本研究首先对玉簪叶片的发育阶段进行了划分,统一了取样时机,然后以幼嫩且未完全展开的玉簪叶束为供试材料,采用专用试剂盒等方法,对8个玉簪品种进行了总糖、总酚、粗蛋白、粗纤维等共计21个指标的测定,最后将所得数据进行隶属函数分析,对8个玉簪品种的营养和品质性状进行了综合评价。测试结果显示,8个玉簪品种在多个营养或品质指标中表现突出,比如全钾、全钙含量、总糖、类黄酮含量等指标最高,含量分别达到(2.84±0.04)mg·kg-1、(3.33±0.06)g·kg-1、(24.60±0.43)mg·g-1、(3.80±0.13)mg·g-1,与莴笋、苹果等比较,含量相当或较高,但维生素C和E含量最高只有(62.68±0.88)μg·g-1和(5.77±0.33)μg·g-1,相比叶用生菜含量较低;隶属函数法综合评价结果表明,德尔塔黎明、镶边的街道、皇标3个品种排名前三,隶属函数平均值较高。本研究对8个玉簪品种的食用前景进行了初步评价,为今后我国观赏食用兼用型玉簪品种的选育、开发推广等工作提供了重要的数据参考。
玉簪属(Hosta)植物属于天门冬科植物, 是我国传统名花。我国有2个原生种, 分别为玉簪(Hosta plantaginea)和紫萼(Hosta ventricosa)[1]。韩国和日本也有分布, 其中韩国产大约11个种和变种[2], 日本产大约18个种和7个变种[3]。有研究认为, 中国两个玉簪属植物200多年前传播到欧洲, 开始了玉簪在花园中的大量应用[4]。据美国玉簪协会的初步统计, 目前玉簪属园艺品种至少有3 000多个, 成为世界范围内重要的宿根花卉。
近年来, 围绕玉簪的栽培、育种、起源、生理生化等内容, 国内外学者开展了大量研究。Kim等[5]研究了在不同温度下一种韩国玉簪(Hosta yingeri)的种子萌发情况, 认为该种种子萌发对温度不敏感, 而其4周幼苗在长日照条件下需要至少26周才能达到50%的开花量。Kanazawa等[6]通过研究认为, 圆叶玉簪(Hosta sieboldiana)的种子降低含水量会延长其寿命, 而含水量低至5%~10%的种子, 在-20 ℃条件下贮藏比5 ℃更容易失去活力。Liu等[7]对46个玉簪种和品种进行了花器官花香成分测定, 认为芳樟醇类物质是影响玉簪花香的主要成分, 而各类花器官中芳香物质的种类是一样的, 只是在含量上略有差异。在栽培应用方面; Liu等[8, 9]对已搜集到的玉簪品种的叶片多样性及光照水平对玉簪生长和观赏性状的影响做了比较研究, 对杂交育种和栽培应用奠定了基础。此外, 玉簪原产地的种质调查和品种起源有了新的进展, Shimasaki等[10]阐明了日本四国岛上原种玉簪的具体分布位置, Lee等[11]在叶绿体DNA水平对31个玉簪品种进行了分析, 探讨了它们的亲本起源。
国内科研机构则对与玉簪属植物的抗逆生理和栽培生理问题进行了大量研究, 夏宜平等[12]较早对花叶玉簪嵌合体的生理机制进行了研究, 从细胞学水平解析了花叶的形成机理。张金政等[13]对氮素水平和光强互作在玉簪生长发育上的影响进行了研究, 得出了同一光照强度下, 玉簪生长发育最大量的氮素需求量。张金政等[14]对东北玉簪和紫萼的抗旱能力进行了评价, 各种生理指标综合分析认为东北玉簪抗旱能力强于紫萼。李金鹏等[15]对彩叶玉簪的光强需求进行了研究, 认为要保持彩叶玉簪的最佳生长和观赏状态, 30%~50%的透光率最有效。罗良旭等[16]在酸雨和镉污染条件下对紫萼的生理生态响应进行了研究, 该工作对复合污染地区的生态修复提供了参考价值。
综上所述, 国内外对玉簪属植物的研究广泛, 从植物学到园艺学领域都有涉及, 但是对于玉簪作为食品开发的研究较少, 而玉簪属植物在日本、韩国有着悠久的食用历史[17, 18, 19], 在我国却几乎是空白。为了拓宽玉簪的经济价值, 本研究从营养学领域经常涉及的指标着手, 开展了8个玉簪品种的营养测定和评价, 以期通过数据的分析, 为我国开发景观、食用功效俱佳的多用途玉簪品种提供借鉴和参考。
本研究所用植物材料为杂交园艺品种, 全部购自浙江虹越花卉有限公司, 4年生组培苗, 起垄地栽, 垄高15 cm, 株行距50 cm× 50 cm。萌芽至取样阶段全光照。选取生长健壮, 无病虫害, 生长发育阶段属于图1所示第3阶段, 即叶片刚刚外展时从基部割取, 生化指标均采用3次重复, 干冰保存备用。玉簪生长第3阶段株高测量采用5次重复。
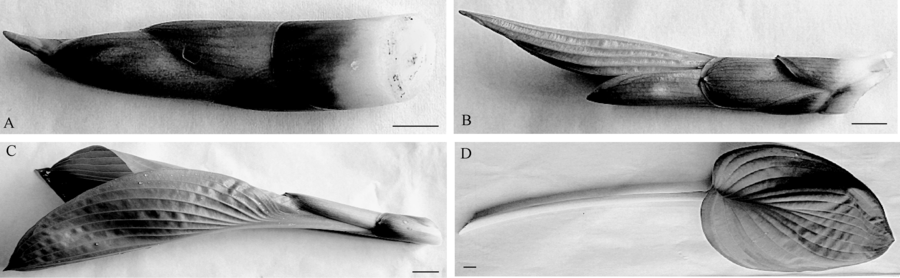 | 图1 玉簪叶片的不同发育阶段A为标尺=0.75 cm; B为标尺=0.55 cm; C为标尺=0.43 cm; D为标尺=0.19 cm |
对总共21个生化指标进行了测试, 均采用苏州科铭生物技术有限公司生产的试剂或试剂盒, 其中总糖含量采用3, 5-二硝基水杨酸法(试剂或试剂盒编号ZT-1-Y, 下同), 可溶性糖含量采用蒽酮比色法(KT-1-Y), 淀粉含量采用蒽酮比色法(DF-1-Y), 单宁含量采用磷钼酸比色法(DN-1-Y), 总酸含量采用滴定法(ZS-1-Y), 类胡萝卜素含量采用可见分光光度法(CAR-1-G), 水分含量采用烘箱干燥法(SF-1-Y), 总酚含量采用钨钼酸比色法(TP-1-G), 类黄酮含量采用铝离子比色法(LHT-1-G), 氨基酸含量采用茚三酮比色法(AA-1-W), 全氮含量采用凯氏定氮法(PQD-1-Z), 全钾含量采用火焰分光光度法(PQJ-1-Z), 全钙含量采用火焰分光光度法(PQG-1-Z), 粗纤维含量采用重量法(CXW-1-T), 抗氧化能力测试(T-AOC)采用血浆铁还原能力法(ferric reducing ability of plasma, FRAP)(TAOC-1-G)。葡萄糖、果糖、蔗糖均含量采用高效液相色谱法(PT-4-Y, GT-4-Y, ZT-4-Y), 包括样品前处理:称取约0.2 g组织, 加入1 mL超纯水, 匀浆, 倒入2 mL EP管中, 超声浸提30 min, 8 000 g 4 ℃离心10 min, 取上清液, 调pH值至5~9, 水系针头式过滤器过滤后待测; HPLC液相条件:Rigol L3000高效液相色谱仪, 糖柱(250 mm× 4.6 mm, 5 μ m), 示差检测器, 流动相:水, 进样量10 μ L, 流速0.6 mL· min-1, 柱温80 ℃, 走样时间为40 min, 最后用流动相过柱子, 待基线稳定后开始加样。绿原酸含量采用高效液相色谱法(CZA-4-Z), 方法概括为:称取0.1 g样本, 放入研钵中磨碎, 加入1 mL预冷的1%的磷酸水溶液(需快速研磨, 防止溶剂挥发), 将液体移入EP管内, 超声提取60 min。10 000 g离心10 min, 提取上清液定容到1 mL, 取适量溶液用针头式过滤器过滤于带有内衬管的样品瓶内待测。HPLC液相条件:Rigol L3000高效液相色谱仪, Kromasil C18反相色谱柱(250 mm× 4.6 mm, 5 μ m), 流动相的配制:A:在1 000 mL超纯水中加入10 mL乙酸配置成1%的乙酸水溶液一瓶。B:1 000 mL甲醇一瓶。开启电脑、检测器和泵, 安装上色谱柱, 打开软件, 在方法组中设置进样量10 μ L, 流速1 mL· min-1, 柱温30 ℃, 波长327 nm, 走样时间为60 min, 用流动相过柱子, 待基线稳定后开始加样。维生素C、维生素E含量采用高效液相色谱法(ASA-4-C)、(VE-4-C), 维生素C测试条件:称取0.1 g样本, 放入研钵中磨碎, 加入1 mL预冷的1%磷酸溶液(需快速研磨, 防止溶剂挥发), 将液体移入EP管内, 10 000 g离心10 min, 提取上清液定容到1 mL, 取适量溶液用针头式过滤器过滤于带有内衬管的样品瓶内待测, HPLC液相条件:Rigol L3000高效液相色谱仪, Kromasil C18反相色谱柱(250 mm× 4.6 mm, 5 μ m), 流动相的配制:A:在1 000 mL超纯水中加入1 mL乙酸配置成0.1%的乙酸水溶液一瓶; B:1 000 ml甲醇一瓶。开启电脑、检测器和泵, 安装上色谱柱, 打开软件, 在方法组中设置进样量10 μ L, 流速1 mL· min-1, 柱温25 ℃, 波长245 nm, 走样时间为15 min, 用流动相过柱子, 待基线稳定后开始加样。维生素E的测试方法为:称取1 g样本(分两管EP管), 各加入1 mL乙醇(需快速研磨, 防止溶剂挥发), 整个过程注意低温、避光、迅速。将液体转移入EP管内, 用乙醇补足提取液至1.5 mL。在50 ℃金属浴中, 振荡提取30 min, 取下冷却至室温, 8 000 g离心10 min, 合并两次上清液至10 mL的EP管中。向10 mL的EP管中加入1 mL 10% NaCl水溶液, 正己烷5 mL, 快速混匀, 漩涡振荡1 min, 静置分层, 取正己烷层并转移到新EP管中, 剩下的液体再重复上述步骤, 合并2次正己烷层, 氮吹吹干, 用0.5 mL流动相复溶, 用针头式过滤器过滤于带有内衬管的样品瓶内待测, HPLC液相条件:Rigol L3000高效液相色谱仪, YMC Carotenoid色谱柱(250 mm× 4.6 mml.D. S-5 μ m), 流动相的配制:在950 mL超甲醇中加入50 mL乙酸乙酯配置成体积比95:5的溶液一瓶。开启电脑、检测器和泵, 安装上色谱柱, 打开软件, 在方法组中设置进样量10 μ L, 流速1 mL· min-1, 柱温25 ℃, 波长275 nm, 走样时间为20 min, 用流动相过柱子, 待基线稳定后开始加样。具体操作样按照试剂或试剂盒使用说明。
本测试所用到的主要仪器有全波长酶标仪Multiskan GO 1510(Thermo, 芬兰), 鼓风干燥机DHG-9035A(慧泰仪器, 中国), 消解仪S402(海能仪器, 中国), 离心机MIRCOCL 17R(Thermo, 德国), 火焰光度计FP6431(仪电分析仪器, 中国), 天平AX205DR(METTLER TOLEDO, 瑞士), 紫外可见分光光度计UV-3000(MAPADA, 中国), 高效液相色谱仪Rigol L3000(普源精电仪器, 中国)等。
采用Excel 2003和SPSS 10.0软件进行试验数据处理分析, 用Duncan新复极差法进行显著性分析。采用模糊数学的隶属函数法综合分析不同品种玉簪营养品质差异[20]。
为了更好地区分不同玉簪品种的生长发育阶段, 将玉簪的叶片发育阶段分为4个阶段。取样时期为第3阶段, 即叶束刚刚展开之际。将该发育阶段的玉簪任选5束进行高度测量, 白边波叶、优雅、德尔塔黎明、巨无霸、镶边的街道、春色、皇标、金叶的叶束高度分别为(15.84± 0.72)cm、(19.62± 1.54)cm、(18.78± 1.05)cm、(24.86± 1.47)cm、(23.80± 0.76)cm、(15.46± 1.01)cm、(26.38± 0.79)cm和(14.92± 0.55)cm。8个玉簪品种在发育第3阶段的高度从大到小依次为:皇标> 巨无霸> 镶边的街道> 优雅> 德尔塔黎明> 白边波叶> 春色> 金叶。同一发育阶段的叶束高度, 间接反映了该品种的长势和生物量大小。
对8个玉簪品种的水分含量、粗蛋白含量、粗纤维含量进行了测定, 结果如表1。结果显示, 8个玉簪品种的水分含量均在0.86 g· g-1以上, 品种巨无霸的水分含量甚至达到(0.90± 0.02)g· g-1, 为8个品种中最高值。各品种的粗蛋白含量差异较大, 数值最高者来自品种德尔塔黎明, 含量为(3.94± 0.07)mg· g-1; 含量最低者来自品种巨无霸, 含量为(1.08± 0.11)mg· g-1, 相差近4倍。粗纤维含量最高者为品种白边波叶, 数值为(12.7± 0.1)g· kg-1; 含量最低者为品种皇标, 数值为(6.8± 0.7)g· kg-1, 说明如果作为蔬菜食用, 皇标的口感较其他品种细腻、不粗糙。
| 表1 不同玉簪品种水分含量、粗蛋白、粗纤维的比较 |
对8个玉簪品种的糖类成分进行了测定, 结果见表2。所测玉簪品种中, 白边波叶的淀粉含量最低, 为(0.35± 0.03)mg· g-1; 春色的淀粉含量最高, 为(0.42± 0.02)mg· g-1, 其余品种间淀粉含量差异不显著。总糖含量最高者为皇标, 达到(24.60± 0.43)mg· g-1; 含量最低者为巨无霸, 仅为(18.77± 0.72)mg· g-1。从可溶性糖含量来看, 皇标含量最高, 其次为春色、德尔塔黎明; 从蔗糖和果糖含量来看, 皇标、德尔塔黎明分别为含量最高者, 如果可食用, 二者口感应较佳。
| 表2 不同玉簪品种淀粉、总糖、可溶性糖、蔗糖、葡萄糖、果糖含量测定 |
表3结果显示, 8个玉簪品种中, 德尔塔黎明的总酚含量远高于其他品种, 其次为金叶和优雅, 但均与德尔塔黎明存在显著差异; 该指标含量最低者为巨无霸。镶边的街道的类黄酮含量最高, 其次为白边波叶和金叶, 但三品种间数值差异不显著; 含量最低者为优雅, 其数值与上述三个品种存在显著差异。品种皇标的类胡萝卜素含量远高于其他品种, 甚至比排序第二的镶边的街道和第三的春色数值之和还要高, 且三者数值存在显著性差异; 该指标含量最低者为优雅。德尔塔黎明的单宁含量在8个品种中最高, 其次为皇标, 但二者数值存在显著差异; 该指标含量最低者为金叶, 数值与其他品种也存在显著差异。金叶的氨基酸含量远高于其他品种, 含量次之的春色与金叶相比也存在显著差异; 氨基酸含量最低的品种为优雅, 与最高品种的数值相差近5倍。总酸指标中, 德尔塔黎明含量最高, 与含量次之的皇标比较, 数值差异不显著, 但与并列第三名的白边波叶和优雅比较, 差异显著; 8个品种中总酸含量最低者为巨无霸。
| 表3 不同玉簪品种总酚、类黄酮、类胡萝卜素、单宁、总氨基酸、总酸含量的比较 |
本研究测定了两种矿物质元素的含量, 其中总钾含量最高的品种为镶边的街道, 其次为德尔塔黎明, 二者数值差异不显著; 该指标含量最低的品种为巨无霸, 与上述品种差异显著。而总钙含量最高的品种为金叶, 其次为皇标, 二者数值差异也不显著; 含量最低者为白边波叶, 与上述两品种差异显著。绿原酸含量最高的品种为镶边的街道, 其次为皇标, 二者存在显著性差异; 该指标含量最低者为金叶和春色, 二者差异不显著, 但与其他品种比较, 均存在显著性差异。品种金叶的维生素E含量最高, 其次为皇标, 二者数值存在显著性差异; 含量最低的品种为优雅。维生素C含量最高的品种为德尔塔黎明和优雅, 二者数值差异不显著; 而含量最低的品种为金叶, 最高和最低含量数值之间存在近6倍的差异。
为了分析不同玉簪品种间抗氧化能力的高低, 本研究采用血浆铁还原能力法(FRAP)对8个品种玉簪的总抗氧化能力进行了初步分析, 白边波叶、春色、德尔塔黎明、皇标、金叶、巨无霸、镶边的街道、优雅的总抗氧化能力为(31.12± 1.82)U· g-1、(32.76± 0.27)U· g-1、(51.72± 0.43)U· g-1、(32.45± 0.71)U· g-1、(34.95± 0.65)U· g-1、(40.79± 0.57)U· g-1、(35.81± 0.61)U· g-1和(54.18± 0.93)U· g-1。数据显示, 在测定的品种中, 优雅的抗氧化能力最强, 其次为德尔塔黎明, 二者数值存在显著性差异; 该指标数值最低者为白边波叶, 数值排名为优雅> 德尔塔黎明> 巨无霸> 镶边的街道> 金叶> 春色> 皇标> 白边波叶。
采用隶属函数法对8个玉簪品种涉及营养、风味、口感等的主要指标进行了综合评价, 各指标赋予相同的权重值, 并且均采用隶属函数公示计算其函数值, 以综合评价供试品种的开发利用价值, 结果见表4。数据显示, 8个玉簪品种的隶属函数值平均值排名, 德尔塔黎明位居首位, 其在总酚、单宁、粗蛋白、维生素C含量方面表现最为突出, 其次为镶边的街道, 该品种在类黄酮、全钾、绿原酸含量方面表现突出; 而排名最后者为巨无霸, 该品种只在含水量方面表现突出。综合评价结果由高到低依次为德尔塔黎明> 镶边的街道> 皇标> 金叶≥ 春色> 优雅> 白边波叶> 巨无霸。
| 表4 隶属函数法对不同玉簪品种进行综合评价分析 |
玉簪是我国传统名花, 也是一味历史悠久的中药材, 虽然栽培年代久远, 但是原产种类只有2种, 园艺品种多为进口。Chung等[2, 3]的研究认为, 韩国和日本拥有较多的玉簪原种, 并且具有较长的食用历史。但是现有文献并未有系统研究玉簪营养成分和品质的相关报道。本研究选取8个玉簪品种, 对其营养、品质相关的21个生理生化指标和1个形态学指标进行了测定分析, 其中叶束发育阶段的划分为本研究取样时机确定了标准, 也为今后同类研究提供了重要参考。粗纤维被认为是食品中重要的营养物质, 并且是直接影响食品口感的成分[21]。本研究中8个玉簪品种粗纤维含量差异较大, 综合评价分析前三位中, 只有镶边的街道粗纤维含量较高, 德尔塔黎明和皇标含量都较低, 口感较细腻。
一般认为, 糖既是果蔬品质和风味物质的主要成分[22], 也是植物抗逆生理中重要的调节物质[23]。本研究针对总糖、淀粉、总可溶性糖以及3种重要甜味糖进行了含量测定。8个玉簪品种, 总糖含量最高值为(24.60± 0.43)mg· g-1, 而有报道指在自然地力条件下, 莴笋叶片的总糖含量最高为(22.3± 0.13)mg· g-1[24]; 蔗糖、葡萄糖、果糖最高含量分别为(6.55± 0.03)mg· g-1、(2.85± 0.04)mg· g-1、(3.74± 0.34)mg· g-1, 而莴笋叶片上述指标含量均略高于玉簪品种[24], 可见部分玉簪品种具备一定的食用开发前景。类黄酮是重要的生物次生代谢物质, 在产品色泽、风味、提高人体生理机能等方面具有重要作用[25]。本研究中品种镶边的街道的类黄酮含量高达(3.8± 0.13)mg· g-1, 而参试品种中含量最低者也有(2.28± 0.23)mg· g-1, 而具有高营养和保健价值的红肉苹果的类黄酮含量为3.0 mg· g-1左右[26], 可见玉簪叶片中类黄酮含量较高。单宁是植物体内一种多酚类物质, 果蔬中含量过多会导致口感苦涩, 但是该物质对抗氧化等生理功能具有重要作用[27], 含量适中则对品质提升具有积极意义。本研究中测定的8个玉簪品种中, 单宁含量在102~157 μ g· g-1, 远低于前人测定的不同柿品种和麻笋等果蔬中的含量[28, 29], 因而不会显著影响口感。
本研究中, 8个玉簪品种全钾的含量在1.71~2.84 mg· g-1, 而前人研究结果显示, 常见蔬菜莴笋、菠菜、大白菜等全钾的含量在1.50~2.20 mg· g-1[30]; 本研究中全钙的含量在2.42~3.33 mg· g-1, 前人研究指莴笋、西芹、菠菜、甘蓝等新鲜蔬菜全钙含量都不超过2.0 mg· g-1 [30]。通过比较, 可见玉簪幼嫩叶束在全钾、全钙含量两项指标上同样具备新型蔬菜开发潜力。但8个玉簪品种的维生素C含量均低于100 μ g· g-1, 明显低于常见蔬菜生菜等的含量[31]; 而维生素E的含量也低于多数水果, 只略高于菠萝中的含量[32], 说明玉簪幼嫩叶束中维生素含量偏低。
隶属函数分析法是一种简单但实用的综合评价测试指标的方法[20]。本研究采用此方法对不同玉簪主要营养指标和抗氧化性能指标等进行了综合测评, 按照数值高低得出, 德尔塔黎明、镶边的街道、皇标三个品种在综合得分中位列前三位, 具备一定的营养优势。但单宁含量和粗纤维含量对玉簪嫩叶的口感和营养功效等的影响尚未明确, 故而本方法未对这2个指标进行反隶属函数分析。近年来, 在牡丹[33]、芍药[34]等观赏植物上进行深加工, 提高产品附加值的研究取得了重要进展, 今后将对8个玉簪品种的有毒有害成分进行测定分析, 以期为其作为果蔬的食用性开发提供理论参考。
(责任编辑:张 韵)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|