

{kind=link}
{kind=link}
棘胸蛙虹彩病毒疫苗制备及免疫技术
[胡霭臻a, b  , 郑善坚
, 郑善坚a, b, * , 林威丞a , 金米雪a , 谢梦佳a , 包伊鹏a , 李柯a ]
, 郑善坚, 林威丞|
|


作者简介:胡霭臻(1995—),女,浙江玉环人,硕士,研究方向为水生动物营养及健康养殖,E-mail:1239762918@qq.com。
对棘胸蛙虹彩病毒(QSIV)进行疫苗制备,通过细胞盲传培养、无菌检验和PCR检测,确定0.1% β-丙内酯4 ℃灭活72 h、0.2%福尔马林37 ℃灭活72 h、1%二乙烯亚胺37 ℃灭活48 h后制备的疫苗安全有效。将制备疫苗对棘胸蛙进行体表喷雾和肌肉注射免疫后,进行血清抗体效价检测和免疫保护力测定。结果表明,注射0.2 mL疫苗组(病毒滴度为1×107 TCID50·mL-1)棘胸蛙21 d血清中和抗体效价最高达到380.2,相对免疫保护率达到80%,为最适的免疫途径。60 min喷雾组28 d时的最高抗体效价为320,相对免疫保护率为60%,可作为快速无损伤免疫的有效途径。
棘胸蛙(Quasipaa spinosa), 又名石蛙, 是我国棘蛙属中分布最广的物种之一。棘胸蛙主要分布于云南、贵州、安徽、江苏、浙江、江西、湖北、湖南、福建、广东、广西和香港等地[1]。20世纪80年代以后, 我国棘胸蛙产业已初具规模, 并且呈现快速发展的趋势[2]。
蛙病毒属病毒宿主广泛, 可以感染爬行类、鱼类和两栖类, 大部分病毒对宿主都有较强的致病性和致死性[3, 4]。2016年本实验室从浙江丽水某养殖场的棘胸蛙上分离到虹彩病毒科、蛙病毒属病毒, 暂命名为棘胸蛙虹彩病毒(QSIV)。感染该病毒的蛙腹部发红, 头部、四肢皮肤溃烂, 死亡率高达90%以上。免疫防治被认为是目前防治蛙等水生动物病毒性疾病的最有效、最安全和最有潜力的方法之一[5], 本文就棘胸蛙虹彩病毒的疫苗制备及免疫方法进行研究。
棘胸蛙虹彩病毒(QSIV)为本实验室提供; 实验用棘胸蛙来自绿谷清泉石蛙养殖合作社, 平均规格(52± 3.5)g; 病毒DNA提取试剂盒购自TIANGEN公司; EPC细胞由浙江省淡水水产研究所惠赠; ELISA试剂盒、M119培养基(Gbico, 11150059)、胎牛血清(BI, 0010316)、β -丙内酯(索莱宝公司, 20170711)、二乙烯亚胺(Sigma, 8044)购自江苏凯基生物公司。
选择生长良好的EPC细胞接种棘胸蛙虹彩病毒, 23 ℃培养箱中培养, 待细胞脱落75%以上时收集病毒, -20 ℃中保存[6]。将所有制得的病毒液混合, 按Reed-Muench法计算病毒滴度[7]。将病毒液稀释至1× 108 TCID50· mL-1, 作为制备疫苗的原材料。
1.3.1 疫苗的制备
根据文献[8], 将收集的病毒稀释液分别用终浓度为0.025%、0.050%、0.100%、0.200%的β -丙内酯(BPL)、福尔马林, 终浓度为0.5%、1.0%、2.0%和3.0%二乙烯亚胺(BEI)灭活剂灭活。BPL灭活温度为4 ℃, 福尔马林、BEI灭活温度为37 ℃; 灭活时间分别为24、48、72、96 h。将制得疫苗暂存在4 ℃冰箱中。
1.3.2 疫苗的安全性检验
将所制得的疫苗分别接种于EPC细胞, 设置2个重复; 同时, 设置接种棘胸蛙虹彩病毒的阳性对照组和灭菌纯水的空白对照组。将未出现细胞病变效应的细胞冻融3次后, 在EPC细胞上盲传3代, 每代培养7 d, 逐日观察细胞病变情况[9]。
1.3.3 疫苗的无菌检验
将所制得的疫苗分别接种于LB固体培养基, 37 ℃培养3 d, 观察培养基中有无细菌生长。
1.3.4 疫苗的PCR检测
根据病毒DNA提取试剂盒提取疫苗和病毒核酸, 以蒸馏水为空白对照进行PCR扩增。PCR条件为预变性95 ℃ 5 min; 变性95 ℃ 40 s; 退火52 ℃ 40 s、延伸72 ℃ 40 s, 30个循环; 再延伸72 ℃ 10 min。反应结束后, 琼脂糖凝胶电泳观察扩增结果[10, 11]。
将棘胸蛙随机均分为6组, 每组20只。3组为注射组, A、B组分别肌肉注射0.2 mL 1× 107 TCID50· mL-1、1× 108 TCID50· mL-1的疫苗, C组注射等剂量的PBS。3组为喷雾组, D、E组分别用喷壶将0.2 mL 1× 107 TCID50· mL-1疫苗喷至雾蛙皮肤, D组喷雾1次, E组每隔10 min喷雾1次, 共喷雾5次, F组喷纯水; 喷雾结束10 min后放回养殖池。免疫后每隔7 d心脏采血1次, 至第28 d共采血4次。将血样室温静置1 h, 4 ℃冰箱静置24 h, 3 000 r· min-1离心10 min, 取上清备用。
1.5.1 血清抗体效价检测
将疫苗12 000 r· min-1离心15 min, 取上清, 用包被液稀释, 在96孔酶标板各孔中加入稀释的抗原100 μ L, 37 ℃温箱孵育, 2 h后取出甩干各孔包被液, 每孔加入封闭液100 μ L, 37 ℃孵育1 h后甩干封闭液, 用洗涤液洗板2次。将待测血清用洗涤液从1:5倍比稀释至1:1 280, 每个稀释度8个重复, 每孔加入100 μ L不同稀释度的血清, 并设置洗涤液为空白对照。37 ℃培养箱温浴1 h, 洗板4次; 在酶标板各孔加入1:5 000稀释的HRP标记羊抗兔IgG 100 μ L, 37 ℃孵育1 h, 洗板5次;
各孔加入TMB显色液100 μ L, 室温避光静置10 min后加入终止剂终止反应。在405 nm波长下, 测定光密度值。阳性判断标准:检测孔D值/阴性孔D值, > 2.1为阳性, < 1.5为阴性, 1.5~2.1为可疑。以出现阳性孔的最高稀释度作为待检血清效价[12, 13]。
1.5.2 相对免疫保护率检测
免疫后35 d, 选择抗体效价较高表喷雾免疫组和注射免疫组进行攻毒试验, 同时设1组未经免疫的空白对照组。每组6只棘胸蛙, 分别肌肉注射0.2 mL的1× 107 TCID50· mL-1病毒液。攻毒后每天观察并记录感染发病与死亡情况, 连续观察21 d, 计算相对免疫保护率[14]。
不同稀释度的病毒液接种后4 d, 出现明显的细胞病变现象, 单层细胞呈破网状(图1)。共制得300 mL病毒液, 混合后, 采用Reed-Muench法计算病毒滴度, 结果为108.125 TCID50· mL-1。
2.2.1 灭活剂灭活浓度和时间
将不同灭活剂、灭活条件制备的疫苗接种于96孔板中的EPC细胞, 将未出现病变的细胞盲传3代。根据细胞的病变情况, 在每组灭活剂中筛选出最适宜浓度和时间的疫苗, 得出使用0.1% β -丙内酯4 ℃灭活72 h、0.2%福尔马林37 ℃灭活72 h、1%二乙烯亚胺37 ℃灭活48 h的疫苗符合上述灭活要求(表1)。
| 表1 疫苗制备条件筛选 |
2.2.2 疫苗安全性
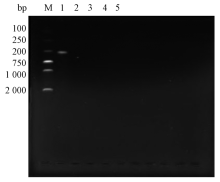
将疫苗接种于普通营养琼脂培养3 d后, 观察到培养基上无细菌生长, 表明疫苗没有细菌污染。将3组最适灭活条件下制备的疫苗, PCR扩增后未检测出虹彩病毒核酸靶基因(图2), 说明0.1% β -丙内酯4 ℃灭活72 h、0.2%福尔马林37 ℃灭活72 h、1%二乙烯亚胺37 ℃灭活48 h能够彻底灭活病毒, 且安全性较好。考虑4 ℃条件下灭活对抗原破坏性小, 确定以0.1%丙内酯4 ℃灭活72 h制备的疫苗进行免疫。
 | 图2 蛙虹彩病毒灭活处理后的PCR鉴定结果M为DNA marker; 1为病毒对照; 2为阴性对照(水); 3为β -丙内酯灭活组; 4为福尔马林灭活组; 5为二乙烯亚胺灭活组 |
2.3.1 血清中和抗体效价
用不同方法对棘胸蛙接种疫苗后, 其血清中和抗体效价相比对照组有显著升高(表2)。注射免疫组在21 d时达到最高的抗体效价, 其中注射0.2 mL 1× 108 TCID50· mL-1疫苗组血清中和抗体效价达到最高的380.2。喷雾免疫组出现最高抗体效价时间晚于注射免疫组, 其中喷雾1次0.2 mL 1× 107 TCID50· mL-1组棘胸蛙血清中抗体效价在28 d时最高, 为160。结果表明, 注射免疫和喷雾免疫都能在一定时间内产生较高的抗体效价, 喷雾免疫需要更长的时间才能发挥较好的免疫作用。
| 表2 注射免疫组血清抗体效价 |
2.3.2 相对免疫保护力
对照组在病毒攻毒感染后的第8 d出现发病症状, 第11 d开始死亡, 21 d时共死亡5只, 死亡率为83.3%。注射免疫和喷雾免疫组在攻毒后第13 d有死亡, 第14 d以后皆无发病死亡情况(表3)。根据公式计算, 注射0.2 mL 1× 108 TCID50· mL-1疫苗组的相对免疫保护率为80%, 喷雾5次组的相对免疫保护率为60%。
| 表3 不同免疫组棘胸蛙死亡情况只 |
病毒灭活是制备疫苗的关键步骤之一。福尔马林是一种最常用的灭活剂, 主要使病毒核酸变性而达到灭活病毒的效果。但是福尔马林对病毒的衣壳蛋白存在破坏作用, 易使病原体蛋白的抗原性降低[15]。β -丙内酯、二乙烯亚胺对病毒具有很强的灭活作用, 能作用于病原体DNA, 改变其结构从而达到灭活目的; 其不直接作用于蛋白, 不破坏病毒的免疫原性, 在维持病毒免疫原性方面优于福尔马林[15, 16, 17, 18, 19, 20]。考虑灭活温度对抗原的结构会造成破坏, 而4 ℃条件对病毒的抗原影响最小, 因此确定0.1% β -丙内酯4 ℃灭活72 h是棘胸蛙虹彩病毒疫苗制备的最适条件。
血清抗体水平的高低与动物机体的免疫保护效果有着密切的关系[21]。采用注射和喷雾的免疫方法对棘胸蛙进行免疫都产生较好的免疫效果。注射0.2 mL 1× 108 TCID50· mL-1疫苗免疫组在21 d时达到最高的血清中和抗体效价380.2, 喷雾1× 107 TCID50· mL-1疫苗5次免疫组在28 d时出现较高的血清中和抗体效价360, 喷雾免疫相比注射免疫发挥免疫保护作用相对滞后。相对免疫保护力检测表明, 注射0.2 mL 1× 108 TCID50· mL-1疫苗免疫组的相对免疫保护率达到80%, 间隔10 min喷雾5次1× 107 TCID50· mL-1疫苗免疫组的相对免疫保护力为60%。因此, 注射0.2 mL 1× 108 TCID50· mL-1棘胸蛙虹彩病毒疫苗后, 免疫保护效果最佳。喷雾疫苗时, 随着喷雾次数增加, 免疫效果进一步增强, 虽然效果上仍不如注射免疫, 但是其操作简便、快速、无损伤, 可作为大规模棘胸蛙虹彩病毒疫苗免疫的有效途径。
鱼类的注射免疫和浸泡免疫已有较多应用[22]。棘胸蛙属于两栖动物, 在免疫系统的发育上要比鱼类高等。两栖类动物的蛙类可以离开水体进行长时间的陆地上生活, 其体表皮肤还富含粘液腺和淋巴间隙, 兼有皮肤辅助呼吸功能。因此, 对蛙类采取体表喷雾免疫, 可以通过皮肤进行疫苗吸收, 刺激机体产生相应免疫作用。研究结果表明, 采用注射免疫或喷雾免疫都能对棘胸蛙虹彩病毒产生较好的免疫保护效果, 为今后蛙病疫苗的开发研究与应用提供良好前景。
(责任编辑:万 晶)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|