

{kind=link}
{kind=link}
南瓜未授粉子房和胚珠离体培养影响因素
[张蕾琛 , 孙楠, 王迎儿, 应泉盛, 严蕾艳, 黄芸萍, 王毓洪, 王洁
, 孙楠, 王迎儿, 应泉盛, 严蕾艳, 黄芸萍, 王毓洪, 王洁* ]
, 孙楠, 王迎儿, 应泉盛, 严蕾艳, 黄芸萍, 王毓洪, 王洁]
 |


作者简介:张蕾琛(1988—),女,浙江象山人,助理研究员,硕士,从事瓜类砧木研究工作,E-mail:sallyzhang910@126.com。
以11个南瓜品种为材料,以未受精子房和胚珠为外植体,研究基因型、激素种类、外植体培养方式和高温预处理时间对胚状体诱导的影响。结果表明,西葫芦材料X2在MS+4.00 mg·L-1 2,4-D+0.5 mg·L-1 6-BA+0.5 mg·L-1 NAA的培养条件下诱导胚状体效率最高,出胚率达15%;子房横切片和胚珠培养对胚珠膨大转绿及胚状体诱导优于子房纵切片培养;外植体在35 ℃高温条件下预处理5 d有利于胚珠的膨大和转绿。
培育单倍体植株在遗传学基础研究和植物育种中具有重要意义。1922年, 意大利的植物学家Blakeslee首次在曼陀罗中发现了自然产生的单倍体植株[1], 引起了植物学家和育种家的广泛关注。随后, 植物学家开始了离体诱导单倍体植株再生的研究。离体雌核发育是指通过离体培养未受精子房或胚珠, 使子房中大孢子或雌配子体向孢子体途径转变, 从而产生单倍体或双单倍体植株的过程[2]。相对于离体雄核发育, 离体雌核发育的研究历史较短, 且由于植物本身生理因素和技术等方面的原因, 利用离体雌核培养获得单倍体植株的技术体系并不完善和成功。但离体雌核发育具有其独特的价值, 尤其在雄核发育诱导单倍体尚未成功或诱导率太低, 白化苗率高, 雄性不育植株及雌雄异株等植物中, 离体雌核发育是获得优良单倍体植株的唯一可行途径[3, 4]。此外, 在某些材料的离体雄核培养中, 植株表现明显的性状变异和倍性变异, 而离体雌核培养的后代则较为稳定。因此, 离体雌核发育作为离体雄核发育的补充, 为单倍体诱导提供了另外一条有效的途径[5]。
葫芦科植物的离体雄核培养难度极大且成功率很低, 因此, 离体雌核培养技术已成为获得葫芦科植物单倍体的重要途径[6]。南瓜(Cucurbita moschata)是葫芦科重要的蔬菜作物之一, 拥有“ 世界性蔬菜” 的美称, 具有丰富的营养和保健功能[7], 获得双单倍体纯系植株对于加快南瓜育种进程具有重要意义。国内外对葫芦科植物离体雌核培养的研究已有报道[8, 9, 10, 11], 但鲜见在南瓜中报道, 且受基因型影响较大[12, 13]。本研究以11个砧木型南瓜一代杂交品种的未授粉子房或胚珠为外植体, 对影响南瓜未授粉子房或胚珠离体诱导胚状体的主要因素进行了探索, 为完善南瓜离体雌核培养技术体系及育种应用提供了理论支持。
试验材料均由宁波市农业科学研究院提供的11份砧木型南瓜一代杂交品种, 其中包括2份西葫芦杂交材料(X1、X2)、5份中国南瓜杂交材料(Z1、Z2、Z3、Z4和Z5)、3份印度南瓜杂交材料(Y1、Y2和Y3)和1份印中杂交南瓜材料Y4。于2017— 2018年3月中旬育苗, 4月下旬定植于宁波高新农业技术试验园区。
1.2.1 外植体取样
在开花前1 d进行扎花, 以确保未授粉。取开花当天的未授粉子房作为外植体, 尽量选取自生长健壮的植株。
1.2.2 外植体处理与消毒
用无菌水清洗外植体表面, 用解剖刀除去花瓣和柱头部分。
对11份南瓜材料的外植体进行以下操作:削去表皮, 将子房纵切成4块, 用75%酒精消毒30 s, 无菌水冲洗1次, 用3% NaClO消毒8 min, 用无菌水冲洗6次, 用解剖刀削去子房壁组织, 用接种针剥离出胚珠, 接种在诱导培养基上。
对南瓜材料X1的外植体进行如下3种操作:1)削去表皮, 将子房横切成1~2 mm厚的切片, 用75%酒精消毒30 s, 无菌水冲洗1次, 用3% NaClO消毒8 min, 用无菌水冲洗6次, 接种在诱导培养基上。2)削去表皮, 将子房纵切成1~2 mm厚的切片, 用75%酒精消毒30 s, 无菌水冲洗1次, 用3% NaClO消毒8 min, 用无菌水冲洗6次, 接种在诱导培养基上。3)削去表皮, 将子房纵切成4块, 用75%酒精消毒30 s, 无菌水冲洗1次, 用3% NaClO消毒8 min, 用无菌水冲洗6次, 用解剖刀削去子房壁组织, 用接种针剥离出胚珠, 将其接种在诱导培养基上。
诱导培养基用IM-4, 接种后的外植体在35 ℃黑暗下热激5 d后, 置于光照(3 500 lx)16 h、25 ℃培养箱中培养, 后续观察其外植体处理方式对诱导胚珠膨大、转绿的影响。
1.2.3 诱导培养基及影响因子
诱导培养基是以Murashige and Skoog(1962)作为基本培养基, 包括大量元素、微量元素和维他命。
8组诱导培养基中添加不同浓度的激素, 具体为IM-1(5.00 mg· L-1 2, 4-D)、IM-2(4.00 mg· L-1 2, 4-D+1.00 mg· L-1 6-BA+0.50 mg· L-1 NAA)、IM-3(4.00 mg· L-1 2, 4-D+1.00 mg· L-1 6-BA+0.25 mg· L-1 NAA)、IM-4(4.00 mg· L-1 2, 4-D+0.50 mg· L-1 6-BA+0.50 mg· L-1 NAA)、IM-5(1.00 mg· L-1 2, 4-D)、IM-6(1.00 mg· L-1 2, 4-D+1.00 mg· L-1 KT)、IM-7(1.00 mg· L-1 6-BA+0.10 mg· L-1 NAA)和IM-8(0.04 mg· L-1 TDZ)。
所有培养基中均添加3%的蔗糖和0.8%的琼脂, pH调至5.7~5.8, 在121 ℃高压灭菌20 min。
1.2.4 胚状体的获得
接种后的外植体在35 ℃黑暗下热激5 d后, 置于光照(3 500 lx)16 h、25 ℃培养箱中6周, 之后在无添加激素的MS培养基中培养, 观察胚状体的形成。
南瓜材料X1, 剥取胚珠后接种在IM-4诱导培养基上。对外植体分别进行0、3、5和7 d不同时间长度的35 ℃黑暗下热激处理, 再置于光照(3 500 lx)16 h、25 ℃培养箱中培养, 观察不同条件下胚珠膨大和转绿的比率。
1.2.5 数据分析
热激完成后记录膨大的胚珠数目, 25 ℃光下培养7 d后记录胚珠转绿数; 培养6周后, 记录胚状体个数。计算胚珠膨大率、胚珠转绿率、胚状体诱导率和成苗率。利用SPSS软件对以上各数据进行方差分析和邓肯多重比较分析。
不同基因型供体材料在相同诱导条件下对培养的反应不一致, 对胚状体诱导有不同程度的表现(表1, 图1)。不同基因型对诱导胚珠膨大(P=0.003)、胚珠转绿率(P=0.001)和胚状体诱导率(P=0.001)有显著影响。其中, 胚状体诱导率最高的是西葫芦组合X1(6.9%), 其次是中国南瓜组合Z1(3.8%)和西葫芦组合X2(3.7%)。胚珠膨大率最高的是印度南瓜组合Y2(90.5%), 但其胚珠转绿率和胚状体诱导率都不高。中国南瓜组合Z3、Z4、Z5和印度南瓜组合Y1、Y3, 以及印中杂交组合Y4虽然都有一定程度的胚珠膨大和转绿, 但却未诱导出胚状体。从不同南瓜类型看, 西葫芦总体比中国南瓜和印度南瓜更易诱导出胚状体。
| 表1 不同基因型对胚状体诱导的影响 |
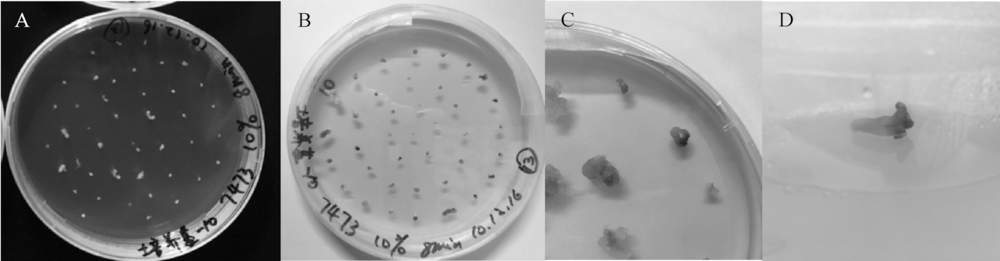 | 图1 南瓜材料Y2未受精胚珠的胚状体诱导及植株再生 A, 从未授粉子房剥离出的胚珠; B, 在诱导培养基上膨大转绿的胚珠; C, 诱导出的胚状体; D, 胚状体初步诱导植株。 |
不同激素对胚状体的诱导结果见表2。不同培养基对胚珠膨大率的影响不显著(P=0.129), 但对胚珠转绿率(P< 0.001)和胚状体诱导率(P=0.001)影响显著。其中, 培养基IM-3胚状体诱导率最高(4.8%); 培养基IM-8胚珠转绿率和胚状体诱导率都较低。同时发现, 外植体在添加2, 4-D的培养基长久培养, 转绿的胚珠会渐渐黄化后发白。
| 表2 不同浓度激素对胚状体诱导的影响 |
不同基因型和不同浓度激素互作效应对胚珠的膨大率(P=0.015)、胚珠转绿率(P=0.018)和胚状体的诱导率(P< 0.001)都有显著影响。由表3可知, 西葫芦材料X2培养基IM-4的诱导胚状体效率最高(15.0%)。
| 表3 不同基因型和不同浓度激素对胚状体诱导的影响 |
本试验通过对南瓜材料X1外植体不同的切片方式来研究其对胚状体诱导的影响。表4表明, 子房纵切法对胚状体的诱导率显著最低, 子房横切片和胚珠剥离对胚珠膨大转绿及胚状体诱导的差异不显著, 但在操作方法上各有利弊。子房横切片操作相对简单, 操作快, 可同时处理的材料数量较多, 但子房切片的厚度和子房除去胚珠的周边组织都会影响胚状体的诱导, 一方面切面会诱导愈伤组织的产生, 一方面诱导出的植株存在可能是胎座细胞诱导产生的可能性。胚珠剥离的方法较繁琐, 且材料本身的胚珠大小对离体胚珠培养也有一定的影响, 相对小的胚珠容易在剥离的时候产生伤口而导致愈伤, 有些则易发白而难以转绿(图2)。
| 表4 外植体不同切片方式对胚状体诱导的影响 |
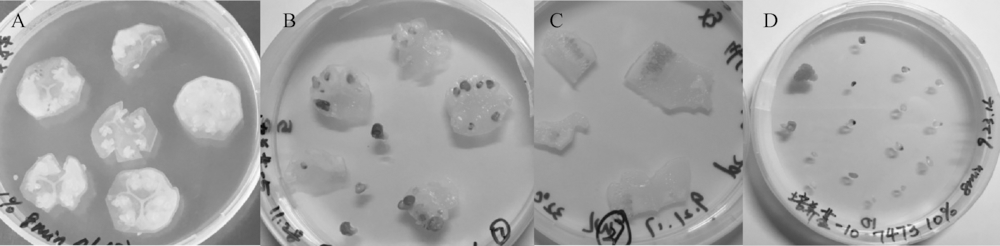 | 图2 外植体不同切片处理诱导胚状体 A, 子房横切片; B, 在诱导培养基上膨大转绿的胚珠; C, 子房纵切片; D, 从未授粉子房剥离出的胚珠。 |
通过对南瓜材料X1外植体不同热激处理时间来研究其对胚状体诱导的影响, 表5显示, 不同热激处理对诱导胚珠膨大(P< 0.001)和胚珠转绿(P=0.001)有显著影响。相比之下, 热激处理可促进胚珠的膨大和转绿, 其中处理5 d的胚珠膨大和转绿的比率最高。
| 表5 外植体不同热激处理时间对胚状体诱导的影响 |
孙守如等[12]研究表明, 从南瓜未授粉子房中剥离出的胚珠在诱导培养基上培养3 d后, 胚珠明显膨大, 颜色变为黄白色。光照培养约18 d后, 形成浅绿色且具光泽的球形胚。大约在诱导培养的20 d, 球形胚发育成典型的心形胚, 并逐渐与母体胚珠组织分离。将心形胚转移至未添加任何激素的培养基上进行成苗培养, 1周后转变为子叶胚。
本研究统计不同因素影响下不同基因型南瓜品种离体雌核的胚珠膨大率、胚珠转绿率和胚状体诱导率发现, 胚珠膨大对诱导胚状体的产生并不产生直接关系, 只有膨大且转绿的胚珠才有可能诱导形成胚状体。因此, 胚珠膨大后进行光照培养形成浅绿色球形胚的时期是诱导产生胚状体的关键时期。
影响植物未受精胚珠孤雌生殖的因素众多。研究发现, 未授粉子房胚状体的诱导率存在明显的基因型依赖性[14], 且胚状体诱导率较高的南瓜品种均为长势较强、细胞分裂较活跃、植株生理状态较好的材料[13]。在本研究中, 11个供试材料中只有5个成功诱导出胚状体, 其他6个材料的胚珠离体培养后, 虽有不同程度的膨大和转绿, 但胚状体诱导率均为0, 说明这些基因型的品种对诱导培养的反应较迟钝。
大量研究表明, 外源激素是影响植物未受精胚珠诱导出胚的决定性因素, 但不同研究者使用的种类和浓度却不完全相同。孙守如等[12]研究发现, 2, 4-D、NAA和6-BA组合有利于南瓜未受精胚珠离体培养胚状体的形成, 出胚效果最好的培养基为MS+1.00 mg· L-1 2, 4-D+0.25 mg· L-1 NAA+0.50 mg· L-1 6-BA, 出胚率达31.1%。而闵子扬等[13]的结果表明, 南瓜未受精子房在MS+4.00 mg· L-1 2, 4-D+0.50 mg· L-1 NAA+1.00 mg· L-1 6-BA和MS+0.04 mg· L-1 TDZ两种培养基上离体培养胚状体的诱导率均较高, 分别为19.8%和20.1%。刘栓桃等[15]认为, 开花当天的西葫芦未受精胚珠在含5%蔗糖和0.8%琼脂的N6基本培养基中添加0.50 mg· L-1NAA和1.00 mg· L-1 2, 4-D最有利于诱导植株再生。程慧等[16]在MS培养基中添加1.00 mg· L-16-BA和0.50 mg· L-1 NAA时, 对西葫芦未受精子房胚状体的诱导效果最好, 诱导率为19.8%。本研究发现, 在MS+4.00 mg· L-1 2, 4-D+1.00 mg· L-1 6-BA+0.25 mg· L-1 NAA的培养条件下出胚率最高, 且不同基因型和不同浓度激素对胚状体的诱导存在互作效应, 西葫芦材料X2在MS+4.00 mg· L-1 2, 4-D+0.5 mg· L-1 6-BA+0.5 mg· L-1 NAA的培养条件下诱导胚状体效率最高(15.0%), 这可能是因为不同基因型材料中的内源激素存在差异, 导致不同基因型材料对不同激素诱导胚状体产生的效率也不尽相同。
本研究发现, 采用子房纵切的方法对胚状体的诱导率较低, 这可能与胎座分布有关, 子房纵切使得胚珠都密集在一个剖面上, 由于切片仅1~2 mm, 导致表面胚珠易被切到而产生愈伤。但子房横切片和胚珠剥离对胚珠膨大转绿及胚状体诱导不存在显著性差异, 这与孙守如等[12]的研究存在不一致。孙守如等[12]认为, 南瓜未受精胚珠离体培养的出胚率(最高为32.2%)高于子房培养的出胚率(最高为20.6%), 这可能是因为剥离胚珠在试验操作上存在一定难度, 操作不当易使胚珠产生伤口而难以转绿形成胚状体。研究表明, 在葫芦科植物中, 对外植体进行35~40 ℃高温预处理可提高其孤雌生殖频率, 但具体机理目前尚不清楚[12]。本研究中, 35 ℃高温预处理5 d, 胚珠膨大转绿和胚状体诱导的比率最高。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|