{kind=link}
{kind=link}
{kind=link}
NaCl和NaHCO3混合盐胁迫对马蔺幼苗生理特性的影响
[袁泽斌1  , 牟昌红
, 牟昌红1 , 王波1 , 孙向丽1, 2, 3, * ]
, 牟昌红]
|
|


作者简介:袁泽斌(1992—),男,山西长治人,硕士研究生,研究方向为园林植物生态修复,E-mail:sudayzb@163.com。
采用室内基质培养法研究NaCl、NaHCO3单盐溶液及其不同配比的混合盐溶液胁迫对马蔺幼苗生理特性的影响。结果表明,幼苗叶片中SPAD值和Pro含量随胁迫时间的延长呈先升后降的趋势,0.30% NaHCO3和0.10% NaCl+0.20% NaHCO3处理的SPAD值在胁迫35 d大幅下降,Pro含量的最大值也高于其他处理;不同盐分组合对可溶性蛋白的影响存在差异;SOD活性在胁迫期间总体呈上升趋势,上升最大的为0.30% NaHCO3、0.10% NaCl+0.20% NaHCO3和0.30% NaCl,各处理的CAT活性在胁迫前期逐渐上升,后期不断下降,POD的活性则为先下降后上升再下降的趋势,升高最多的为0.30% NaHCO3、0.30% NaCl处理。综上认为,NaCl和NaHCO3两种单盐的胁迫在抗氧化酶活性等方面较混合盐影响大且碱性盐胁迫作用强于中性盐,混合盐胁迫中NaHCO3浓度占主导作用。
盐碱土在我国分布广泛, 严重制约着农业生产。我国耕地盐碱化面积超过760万hm2, 占总耕地面积的1/5, 主要分布在东北、华北及西北内陆地区[1]。盐碱土中的离子会抑制植物对养分的吸收[2], 还会提高土壤中重金属的迁移能力[3]。采用植物改良的方式虽周期较长, 却能从根本上改善土壤理化性质, 同时还能产生经济效益, 被认为是最经济持久的方法[4]。
马蔺(Iris lactea var. chinensis)又称马兰、马韭, 为鸢尾科鸢尾属多年生草本宿根植物[5], 其花大色艳, 具有较高的观赏价值, 且根、花、种子均可入药[6]。马蔺根系发达, 具有极强的耐干旱、耐盐碱等特性[7]。中国盐碱地土壤中主要致害离子为Na+、Cl-、HC
供试植物马蔺的种子由江苏省沭阳斗研种业有限公司提供。选取籽实饱满, 种皮完好的马蔺种子, 在50 ℃以下的温水中浸种3 d, 期间每天更换水2~3次, 最后1次换水时把生根粉倒进去一起浸泡10 min, 捞出晾干, 然后播种在盛有石英砂和泥炭的基质中, 覆盖薄膜, 每3 d浇1次水, 保持基质处于湿润状态, 待幼苗长至10 cm时, 移栽至直径10 cm的小花盆中, 并用盐溶液处理, 每个小花盆中盛100 g基质。
1.2.1 幼苗的处理方法
试验于2019年3— 4月在苏州大学建筑学院栽培生理实验室进行。将栽有马蔺幼苗的小花盆放置在规格相同的白色托盘中, 每个托盘中放置相同数量的马蔺幼苗, 以清水处理为空白对照, 其余5个处理分别为0.30% NaCl、0.20% NaCl+0.10% NaHCO3、0.15% NaCl+0.15% NaHCO3、0.10% NaCl+0.20% NaHCO3、0.30% NaHCO3五种盐溶液组合(所有浓度均为质量分数), 分别将配制好的盐溶液以每盆100 mL倒入托盘中, 让马蔺幼苗充分吸收盐溶液, 直至完全吸收。试验过程中, 每7 d取1次样, 并给幼苗浇1次水, 每处理幼苗80棵, 为减小试验误差, 每次取样时随机选取15棵剪取幼苗叶片, 每5棵为1个重复, 每处理3个重复, 共取样5次, 每次取样完后剪取了叶片的幼苗弃用。
1.2.2 生理特性指标测定
SPAD值的测定采用SPAD-502叶绿素仪, Pro含量测定采用磺基水杨酸比色法, 可溶性蛋白含量测定采用考马斯亮蓝法, SOD、POD、CAT酶活性的测定采用苏州科铭生物技术有限公司生产的试剂盒进行测定。
采用Excel 2010软件进行数据的整理与图表的绘制, SPSS 22.0进行数据的统计分析, 运用Duncan’ s新复极差法进行差异显著性检验(P< 0.05)。
SPAD值(绿色度)与叶子的叶绿素含量呈显著正相关, 可对叶片中的叶绿素水平提供可靠估计[9]。表1显示, 通过对盐胁迫后的马蔺幼苗叶片的SPAD值的测定表明, 在胁迫第7天时, 各处理的幼苗叶片叶绿素含量未出现显著差异。随着胁迫时间的延长, 对照处理的幼苗叶片叶绿素含量呈显著递增趋势, 而其他处理均为先增长后降低。在胁迫第21天时, 除对照外, 其余各处理的幼苗叶片叶绿素含量达到最大, 而后呈下降趋势。在胁迫第35天时, 各处理叶绿素含量与对照相比均显著降低, 其中含量最低的为0.10% NaCl+0.20% NaHCO3处理的幼苗叶片, 仅为28.90, 较对照降低29.73%; 其次为0.30%NaHCO3处理的幼苗叶片, 为29.50, 较对照降低28.28%; 混合盐中NaHCO3浓度越大, 幼苗叶片中的叶绿素含量越低。
| 表1 不同盐分组合对马蔺幼苗叶片SPAD值的影响 |
Pro积累与胁迫耐受性有关[10, 11]。Pro被认为是一种渗透调节剂, 可保护细胞免受应激造成的损伤[12, 13]。通过对盐胁迫后不同时间马蔺幼苗叶片中Pro含量的测定(表2)表明, 空白对照组的马蔺幼苗叶片中的Pro含量变化较稳定, 其余各处理随着胁迫时间的延长均呈先增长后下降的趋势。在处理的第7天, 经0.3% NaHCO3溶液处理的幼苗叶片中Pro含量较其他处理已显著上升, 达到5.04 μ g· g-1; 第21天时含量达到最大, 为7.34 μ g· g-1, 为空白对照的1.70倍; 在第35天时, 经0.30% NaCl和0.20% NaCl+0.10% NaHCO3处理的幼苗叶片中的Pro含量较对照显著下降, 0.15% NaCl+0.15% NaHCO3、0.10% NaCl+0.20% NaHCO3、0.30% NaHCO3处理中Pro含量仍显著高于对照组。
| 表2 不同盐分组合对马蔺幼苗叶片Pro含量的影响 |
可溶性蛋白是一种重要的渗透调节物质和营养物质, 其增加和积累能够提高植物细胞的保水能力, 对细胞的生命物质及生物膜起到保护作用, 也是筛选抗逆性的指标之一[14]。通过对盐溶液胁迫后不同时间马蔺幼苗叶片中可溶性蛋白含量的测定(表3)表明, 随着胁迫时间的延长, CK处理的可溶性蛋白含量较稳定, 0.30% NaCl、0.20% NaCl+0.10% NaHCO3、0.15% NaCl+0.15% NaHCO3处理幼苗叶片中的可溶性蛋白含量表现出“ 下降、上升” 的趋势, 而0.10% NaCl+0.20% NaHCO3、0.30% NaHCO3处理则表现为“ 下降、上升、下降” 的趋势。在胁迫的第7天, 可溶性蛋白含量最低的为0.30% NaHCO3处理, 仅为0.238 mg· g-1, 较0.30% NaCl、0.20% NaCl+0.10% NaHCO3处理显著下降; 第21天时, 0.10% NaCl+0.20% NaHCO3、0.30% NaHCO3处理的可溶性蛋白含量较其他处理显著升高, 分别达到0.273和0.285 mg· g-1; 第35天时, 可溶性蛋白含量最高的为0.30% NaCl处理的幼苗叶片, 而0.10% NaCl+0.20% NaHCO3、0.30% NaHCO3处理较其他处理显著降低, 推测原因可能是这两个处理对幼苗的胁迫作用较强, 导致可溶性蛋白逐渐降解。
| 表3 不同盐分组合对马蔺幼苗叶片可溶性蛋白含量的影响 |
2.4.1 SOD酶活性
植物的抗氧化酶系统包括超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)、谷胱甘肽还原酶(GR)等多种酶类, 是植物在遭受逆境胁迫时重要的防御体系[15]。各酶活性之间的平衡可将超氧自由基和H2O2维持在稳定水平[16]。SOD是抗氧化酶系统中的第一道防线[17], 其作用是保护细胞免受超氧化物诱导的氧化胁迫[18]。通过对盐胁迫后不同时间马蔺幼苗叶片中SOD活性的测定(图1)表明, 经各盐胁迫处理的幼苗叶片中的SOD的活性均始终高于空白对照组, 0.30% NaCl、0.20% NaCl+0.10% NaHCO3、0.15% NaCl+0.15% NaHCO3处理随着胁迫时间的延长总体上呈现出不断上升的趋势。在第35天时, SOD活性分别为第7天的1.72、1.56、1.70倍; 在第28天时, 0.10% NaCl+0.20% NaHCO3、0.30% NaHCO3处理的SOD活性达到最大, 其中0.30% NaHCO3处理的活性达到1 455.25 U· g-1, 为对照的1.73倍; 到第35天出现略微下降, 说明在盐胁迫的过程中, 马蔺幼苗可通过提高SOD的活性来保护细胞免受超氧化物的影响。
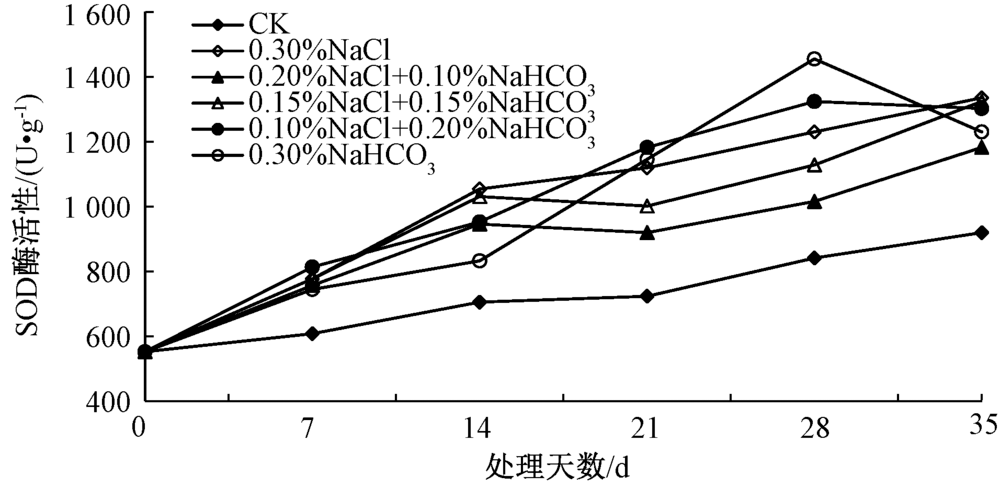 | 图1 不同盐分组合对马蔺幼苗叶片SOD酶活性的影响 |
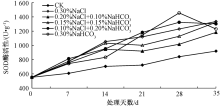
2.4.2 CAT酶活性
CAT广泛存在于植物中, 是一种包含血红素的4聚体酶, 在遭受逆境胁迫时, CAT主要负责清除光呼吸和脂肪酸β 氧化过程产生的H2O2[19]。通过对盐胁迫后的马蔺幼苗叶片中CAT活性的测定(图2)表明, 除空白对照组外, 各处理幼苗叶片中的CAT酶活性都呈现出“ 上升、下降” 的趋势。其中, 0.30%NaCl、0.20%NaCl+0.10%NaHCO3、0.15%NaCl+0.15%NaHCO3处理的CAT酶活性在胁迫第21天时达到最大, CAT活性最低的为0.20% NaCl+0.10% NaHCO3处理, 0.10% NaCl+0.20% NaHCO3、0.30% NaHCO3处理的CAT活性在胁迫第28天时达到最大, 其中0.30% NaHCO3处理的酶活性达到380.97 U· g-1, 为对照的1.31倍; 在第35天时, 各处理的CAT酶活性均较对照小, 可能原因是在胁迫初始阶段, 幼苗体内产生过多的H2O2, CAT活性不断升高以清除H2O2来维持内环境的稳定, 随着胁迫时间的延长, H2O2不断积累, 毒害植物细胞, 致使CAT活性出现急剧下降。
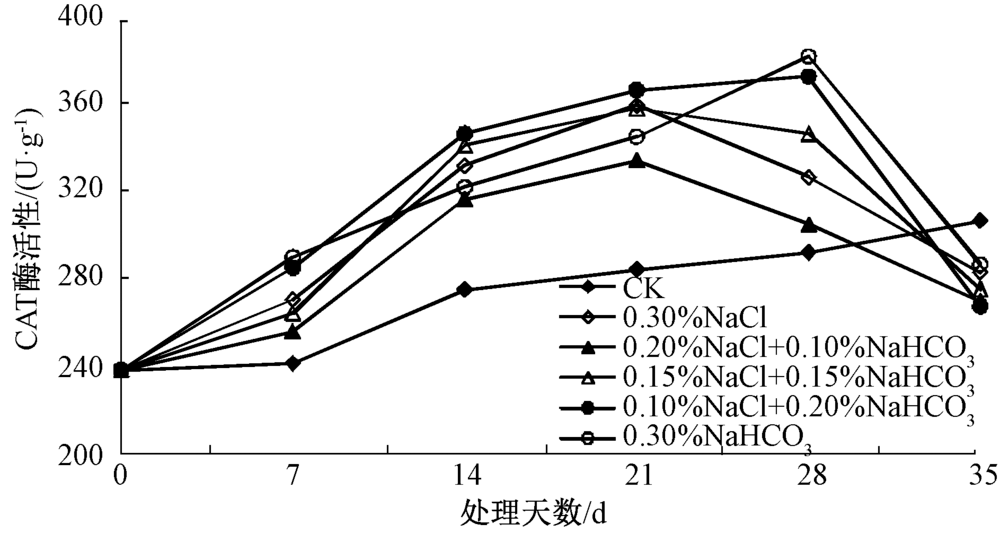 | 图2 不同盐分组合对马蔺幼苗叶片CAT活性的影响 |
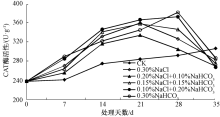
2.4.3 POD酶活性
POD可催化过氧化氢氧化酚类和胺类化合物, 具有消除过氧化氢和酚类、胺类毒性的双重作用[20]。通过对盐胁迫后不同时间马蔺幼苗叶片中POD活性的测定(图3)表明, 在胁迫初期, 各处理幼苗叶片中的POD活性均呈现出略微下降的趋势; 在第21天时, 0.30% NaCl、0.20% NaCl+0.10% NaHCO3、0.15% NaCl+0.15% NaHCO3处理的POD活性急剧升高, 最高的为0.30% NaCl处理, 达到3 666.67 U· g-1, 为空白对照组的2.00倍, 而0.10% NaCl+0.20% NaHCO3、0.30% NaHCO3处理在第28天时酶活性达到最大, 分别为空白对照组的1.73和1.88倍; 在第35天时, 除0.10% NaCl+0.20% NaHCO3处理的POD活性较对照略高外, 其余处理已降至低于空白对照的水平, 原因可能为在初始阶段, 由于CAT活性的升高及在清除H2O2的过程中起到主导作用, 随着H2O2的不断积累, CAT的作用有限, POD活性开始升高, 与CAT共同作用以清除细胞中的H2O2等有毒有害物质, 最终由于盐胁迫作用的持续, 导致POD活性也出现下降趋势。
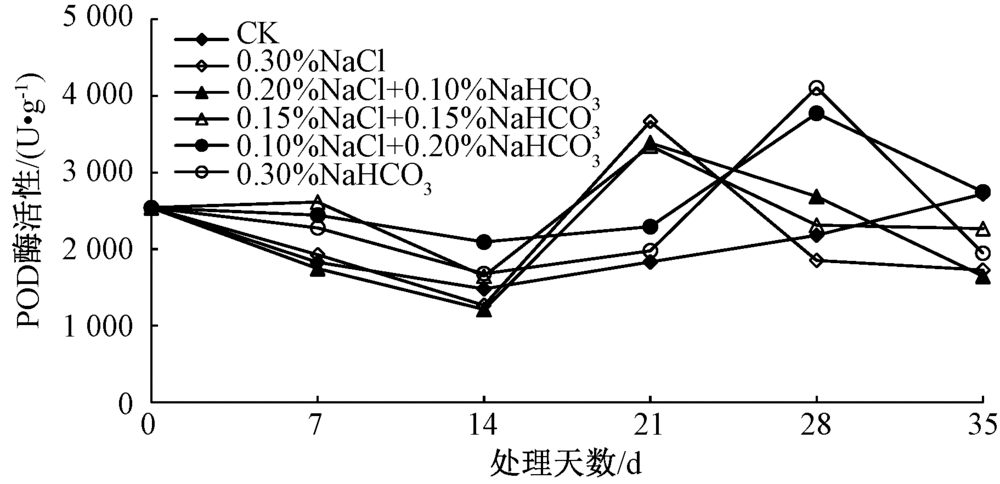 | 图3 不同盐分组合对马蔺幼苗叶片POD活性的影响 |
植物耐盐的生理机制包括逃避盐害和忍受盐害两种方式, 渗透调节以及在抗氧化酶系统下参与的膜系统的保护, 都属于忍受盐害的方式[21]。在盐碱胁迫下, 植物可以通过合成Pro、可溶性蛋白、醇类等有机调节物质来维持细胞内的渗透平衡, 这些物质还能起到稳定细胞质中酶分子活性构象的作用[22, 23]。研究发现, 燕麦[24]、小麦[25]、玉米[26]等农作物在遭受盐胁迫时可积累Pro和可溶性蛋白来维持正常的生命活动。本研究也发现, 在盐胁迫过程中, Pro和可溶性蛋白含量会有所提高, 但随着胁迫时间的延长, Pro含量在胁迫后期有所下降, 这与张明轩等[27, 28]在研究NaCl对马蔺幼苗生长的影响时所得出的结论相同。SPAD值可反映叶片中叶绿素的含量。本研究发现, 在胁迫时间内, 叶片中的叶绿素含量呈先升后降的趋势, 这与刘文瑜等[29, 30]在藜麦、朴树和速生榆上的研究结果一致。植物耐盐的生理机制是不同作用之间相互协作的, 抗氧化酶的保护作用可与渗透调节同时存在。本研究中, 马蔺幼苗在盐胁迫作用下SOD活性会不断升高, CAT活性会随着SOD作用不断产生H2O2而升高, POD的活性则会在CAT清除H2O2能力不足时升高。马晶晶[21]在研究NaCl盐胁迫对马蔺幼苗生长的影响时发现, SOD的活性是先降后升的, 原因可能是, 其处理幼苗所用NaCl浓度为1.5%, 为本研究的5倍, 在高盐胁迫下所表现出的酶活性变化和低盐胁迫下有所差异。
本研究通过用单盐和混合盐对马蔺幼苗进行胁迫发现, 单盐的胁迫作用总体上较混合盐的胁迫作用强, 且碱性盐的胁迫作用较中性盐胁迫作用强。白文波等[31]在研究NaCl和NaHCO3胁迫下马蔺生长与光合特性时发现, 碱性盐特有的高pH会导致植物体内的生理代谢紊乱, 也证明碱性盐胁迫强于中性盐。刘丹等[32]发现, 单盐对狗枣猕猴桃幼苗的胁迫作用强于混合盐, 这与本研究结论一致, 但其研究结果却认为, NaCl胁迫作用强于NaHCO3, 原因可能为狗枣猕猴桃为大型藤本植物, 对NaCl和NaHCO3的响应机制不同。
本研究对马蔺幼苗的耐盐机制做了生理上的初步探究, 马蔺耐盐基因值得深入研究。本研究采用的为基质培养幼苗, 浇灌盐溶液的方式模拟盐碱土, 与真正的盐碱土环境存在着一定的差别, 所以在今后的研究中也要注重在实际中的应用, 为马蔺在盐碱土改良方面的推广提供可靠的依据。
综上所述, 通过本研究得出, 在胁迫35 d中, 各盐溶液处理后的马蔺幼苗叶片的SPAD值和Pro含量均呈现出先上升后下降的趋势, 其中经0.10% NaCl+0.20% NaHCO3和0.30%NaHCO3溶液处理的幼苗叶片的SPAD值在第35天时下降最显著; 在第21天时, Pro的含量在5个处理中也最多, 这两个处理的幼苗叶片中的可溶性蛋白的变化趋势与其他处理也不完全相同, 表现为“ 下降、上升、下降” 的趋势, 而其余处理则为“ 下降、上升” ; 5个处理对抗氧化酶系统中的各种酶活性的影响也存在较大的差别, 在胁迫期内, SOD活性总体呈上升趋势, 仅0.10% NaCl+0.20% NaHCO3和0.30%NaHCO3处理的幼苗叶片中SOD活性在第35天时出现略微下降, CAT和POD活性的变化趋势则大体相同。由此可得, 在单盐中, 碱性盐的胁迫作用较中性盐的胁迫作用强; 混合盐溶液中, 碱性盐的浓度越大, 胁迫就越严重, 除0.10% NaCl+0.20% NaHCO3盐溶液组合外, 单盐的胁迫作用较另外两个盐溶液组合强。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|